雜交種馬鈴薯育種的制作方法
本申請是申請日為2012年04月20日,申請號為201280031056.2,發明名稱為“雜交種馬鈴薯育種”的分案申請。本發明涉及農業領域,尤其涉及新型植物及用于獲得該新型植物的工序。本發明還涉及用于改善農作物的基因組成的方法,且涉及由該方法獲得的植物。本發明進一步涉及用于生產雜交種子的方法,且涉及由該方法生產的雜交種子,且涉及使用本發明的雜交種子生產作物的方法。
背景技術:
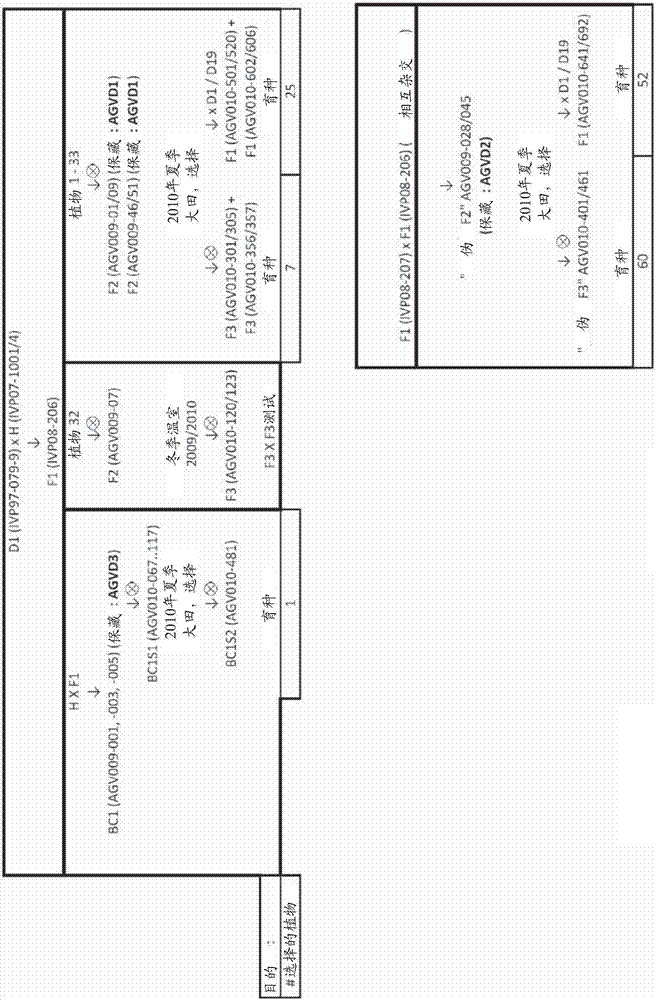
:馬鈴薯(solanumtuberosuml.)是全球糧食系統的基本部分。它是世界上第一位的非谷物糧食商品,在2007年具有達記錄325000000噸的產量。與其它主要的大田作物不同,馬鈴薯是從其它馬鈴薯繁殖生長的。因此,留出每年一部分的作物(從5%至15%的范圍,依賴于收獲的塊莖的質量)用于在下一種植季節中使用。在發展中國家,大部分農民選自且儲存他們自己的種子塊莖。在發達國家,農民更有可能從制定供應商處購買無病的“合格種子”。種用馬鈴薯比谷物或脈沖種子(pulseseed)更難以生產和供應。與玉米的1:400或番茄的1:10000相比,馬鈴薯的種子:收獲量的比率為1:20被認為是好的。因此,與玉米的18kg相比,一公頃可能需要兩頓的種料以最大化可收獲的產物的產量。為了打破休眠,種用馬鈴薯應在它們可以種植之前儲存幾個星期。在儲存過程中的正確條件(如光量、溫度和濕度)對確保良好的“種子”質量起到至關重要的作用。除了不理想的種子:收獲量的比率之外,種用馬鈴薯引起且轉運害蟲(pest)和疾病。這包括(其中包括)晚疫病菌、安第斯馬鈴薯甲蟲、線蟲、塊莖蛾和病毒。后者在田間通過蚜蟲傳遞,且然后在種子被一代接一代地攜帶。這樣的病毒感染可以降低高達20%的產量。因為在主要種子生產區至主要消費品生產區之間的距離遙遠,且各種用馬鈴薯塊莖具有較沉的重量,因此種用馬鈴薯具有高轉運費用。為了滿足現在和將來逐漸增長的世界人口,馬鈴薯產業必須保持增殖以滿足消費者的需要。基礎研究和開發工作致力于使大田種植和收獲及馬鈴薯的加工現代化,且開發經濟有利的馬鈴薯品種。通過馬鈴薯雜交,研究者希望獲得具有用于新鮮消費以及用于產業用途的良好加工、高可溶的固體含量、高產量、對疾病和害蟲具有抗性及能夠適應各種生長地區和條件的期望特征的馬鈴薯。導致產生與上述有利特征結合的馬鈴薯品種的研究主要以經驗為依據。該研究需要投入大量的時間、人力和金錢。馬鈴薯栽培品種的開發可以經常花費八年或更長的時間,然后進行至少五年的培植以得到足以用于商業用途的數量。育種從仔細選擇較好的植物以將最重要的特征并入后代中。由于所有期望的性狀通常不顯現在一世代中,因此育種是連續選擇最佳結合(結合原種的良好性狀)的過程。當理解了馬鈴薯的遺傳學時,能最好理解產生馬鈴薯新型品種的艱苦工作。商品化馬鈴薯具有四倍體的基因組。二倍體的塊莖對于重要的商業應用來說通常太小。此外,四倍體基因組是極端雜合型的,每一基因座均含有多個等位基因。據信主要在二倍體水平上經歷的自交不親和性,及近交衰退是造成維持在馬鈴薯中發現的高遺傳變異性的原因,且據信雜合的等位基因的超顯性(雜種優勢)產生茁壯的植物。在兩個無關的親本之間的雜交所獲得的典型的馬鈴薯后代中,有害的同位基因由此可能在純合性的情況下有助于適合度下降,或在雜合性的情況下有助于增強活力。很明顯,培植者需要大量種群以最大化發現攜帶相對大量的雜合性基因座及少量純合性基因座,同時還顯示出農藝學上所期望的性狀的有益組合的個體的機會。現在的馬鈴薯育種技術依賴于自身是綜合預雜交開發的結果的親本克隆(parentalclone)的受控雜交,在該過程中除了其它技術之外,還應用了特定的技術(如染色體加倍、種胚挽救(embryorescue)及體細胞融合)以將例如野生型和原生型茄屬植物的有益特征引入這些克隆中。然后,使在表型選擇程序之后發現適用于進一步育種的親本材料相互雜交,且得到的不一致的雜交種子被大量播種在溫室中。從成千上萬的個體f1中收獲幼苗塊莖,且保持該幼苗塊莖用于下一年栽培。下一年,來自每一得到的幼苗的單個“種子”塊莖被種植在大田中。必須極其謹慎以避免引入病毒和疾病,因為材料在它幾年后被賣給個體消費者之前僅被無性繁殖(clonally)(植物性)擴增。在第二年后,取出塊莖的樣品進行密度測量和初步油炸測試(preliminaryfrytest)以確定塊莖用于商業用途的適合性。然后,已經從選擇過程中幸存下來至這一點的眾多植物的塊莖被種植三年,用于綜合性更高的系列油炸測試和密度測定。在開發的第四年階段,遞減數量的幸存的選擇以始終擴增的量進行生長,且對其植物進行幾個階段的大田試驗以確定它們對不同生長條件的適合性。最后,具有優秀農業學質量的品種被轉移至其它農場,且“種子”(以塊莖的形式)被增加至商業規模。由于一個“種子”塊莖可產生6至20個之間的收獲的塊莖,該放大的過程可能花費幾年以產生足夠的“種子”。通常,到這個時候,已經在嘗試開發新型和改良的馬鈴薯栽培品種中投入了8年或更長時間進行種植、收獲和測試。為了降低近交衰退,培植者可以引入來自遺傳學上相隔更遠的具有從四倍體至六倍體范圍的多倍性的親本(如從野生型和原生型品種)的新基因。但是,當兩株遺傳學上沒有聯系的馬鈴薯植物雜交時,雜合性水平可以增加,但同時還引入了更加有害的基因。因此,培植者應典型地與更商品化的種質進行雜交以富集含有利等位基因的種群。同時,這樣的多次雜交育種的程序可能進行幾十年,如在每一世代中選擇有利基因型可能已經進行了五年。因此,當前馬鈴薯育種是以經驗為主的活動,特別具有試驗和錯誤的特征。馬鈴薯及其有聯系的野生型品種(結出塊莖的茄屬植物)是最多的遠交交配,因為配子體自交不親和系統阻礙了自受精。自交不親和(si)是被子植物中幾種遺傳機制的通用名稱,si防止了自受精和近親交配。在具有si的植物中,當在植物中產生的花粉粒達到同一植物或具有相似基因型的另一植物的柱頭時,花粉萌發、花粉管生長、胚珠受精和種胚發育在它一個階段處被停止,且隨后不會產生種子。在四倍體馬鈴薯中沒有發現自交不親和。這樣的自交親和的克隆的供應可以促進馬鈴薯的自花傳粉的后代的產生,且由此產生(高)純合的馬鈴薯品系。這樣可以提供極好的機會以在馬鈴薯中開發純合的優秀育種品系。但是,到目前為止,開發具有遺傳固定的農藝學上期望性狀的純合的優秀育種品系還沒有成功,該優秀育種品系將能夠產生遺傳上一致的雜交馬鈴薯種子。提供純合的良種品系被未知的原因阻礙。偶爾遇到的自交親和克隆的自交導致雜合度比理論上所預期的更慢的速度降低。在顯示更高活力、能育性和種子萌發的后代中,雜合度更慢的下降可以是在雜合植物的自交過程中的無意但不可避免的選擇結果。這暗示,如雜合度通過自交降低,能育性和活力也降低,且植物可變得弱小且完全不育。該情況可能由于隱性有害基因的純合配置的建立而變得更壞。這種現象(通常被稱為近親交配衰退)已經大大阻礙了純合馬鈴薯品系的開發,且由此阻礙了一致雜交馬鈴薯種子的產生。由于近交衰退,具有不利于在馬鈴薯中產生純合育種品系的損害。uijtewaal等.(euphytica36(1987)745-753)示出歸因于不育的問題,純合馬鈴薯克隆對于實際育種不是很重要。考慮了開發純合近交品系,但在馬鈴薯中不可能(umaerus,1987,第10屆歐洲馬鈴薯研究協會的會議記錄(proceedingofthe10thtriennialconferenceoftheeuropeanassociationofpotatoresearch),奧爾堡,丹麥,pp72–103,如在almekinders等.2009potatoresearch52:275–293中所引用的)。涉及單倍體加倍的途徑已經長時間推測為是有希望的。但是,直到今天,流行的觀點是雙倍體馬鈴薯中的近親衰退太強大,從而不能得到有活力的純化植物。birhman和hosaka(genome43:495–502(2000))已經建議利用來源于s.chacoense的sli基因用于開發高度純合真實的馬鈴薯種子(tps)品系和進行馬鈴薯雜種優勢育種的可能性。但是,到目前為止,還沒有報道來自該研究的推薦品系的具有良好農藝學性狀(如良好的塊莖產量)的純合品系。準確地說,已經產生的那些純合體不顯示任何農藝學相關的塊莖產量。由于努力改善該作物的產量和質量被近交衰退所阻礙的事實,rommens在2010年(植物的遺傳修飾(geneticmodificationofplants),kempen&jung,編輯在biotechnologyinagricultureandforestry64(1):61-77(2010)中)主張遺傳轉化的途徑。簡單講,認為產生馬鈴薯的真實的育種品系是不可能的。由于這個事實,馬鈴薯育種不可能逃脫基于四倍體雜合體雜交的傳統方案。結果,挑戰被認為是難以克服使各種利用性狀(與新型和加工使用(processinguse)有關)、對病原體和害蟲具有抗性、及在產量上具有改善的其它相關的農藝學性狀與商業上可接受的栽培品種相組合(douches等.1999,cropscience36(6):1544-1552)。本發明的目標是提供用于產生馬鈴薯優秀育種品系和用于產生從中可以生長顯示農藝學上相關塊莖產量的植物的一致的雜交馬鈴薯種子的手段和方法。技術實現要素:本發明人曾試圖通過多條路線實現用于馬鈴薯生殖增殖的純合親本育種品系,但沒有一條路線成功。這些失敗路線包括雄核發育、雌核發育、使單倍體加倍和連續自交。只有使包括來源于s.chacoense的sli基因的自交親和的植物重復自交并使用時,發明人才得到了本發明的產品。現在本發明提供了如在所附權利要求中所引用的方法和植物。本發明在第一方面提供了一種二倍體、能育的、自交親和且基本純合的馬鈴薯品系,該馬鈴薯品系包括具有以每株植物至少200克鮮重的克數表示的平均塊莖產量的植物。具有以每株植物至少200克鮮重的克數表示的平均塊莖產量的植物在本文中也被一般地稱為有活力(vigorous)的植物。在優選實施方案中,所述品系除所述塊莖產量之外還包括至少一個、更優選至少2、3、4、5或6個(附加的)農藝學期望的性狀。優選地,所述農藝學期望性狀選自由昆蟲耐性、線蟲抗性、疾病抗性(包括但并不限于對粉痂病(powderyscab)、絲核菌(rhizoctonia)、銀屑病(silverscurf)、晚疫病菌(phytophthorainfestans)、鏈霉菌(streptomycesspp)導致的瘡痂病(scab)的抗性)、除草劑耐性、耐寒性、耐旱性、耐濕性、耐干腐和濕腐性、耐鹽性及對低溫糖化的抗性所組成的組。在本發明所述馬鈴薯品系的另一優選實施方案中,所述品系的植物成熟時達到枝葉和芽的平均鮮重為單株植物至少500克。枝葉和芽的平均鮮重達到單株植物至少500克的植物在此也被一般地稱為有活力的植物。另一方面,本發明提供一種如上所述的根據本發明的品系的植物。優選地,所述植物是二倍體、能育的、自交親和且基本上純合的植物,且具有以每株植物至少200克鮮重的克數表示的平均塊莖產量。另一方面,本發明提供一種如上所述的植物的種子。所述種子可以是純合的(即表示自交的種子),對親本不同的等位基因來說也可以是雜合的。在一個實施方式中,所述種子優選為一致的雜種馬鈴薯種子。該一致的雜種馬鈴薯種子是如上所述的根據本發明的品系(優選兩個不同品系)中的兩個純合的、自交親和的、能育且有活力的植物之間雜交的結果,其中所述品系包含通過分子標記物檢驗可確定的至少20%的相對的純合的基因座(contrastinghomozygousloci)。另一方面,本發明提供一種用于產生二倍體、能育的、自交親和且基本純合的馬鈴薯品系的方法,所述馬鈴薯品系包括具有以每株植物至少200克鮮重的克數表示的平均塊莖產量的植物,所述方法包括:(a)提供第一馬鈴薯植物,所述第一馬鈴薯植物是二倍體、自交親和的、能育的且基本純合的第一馬鈴薯品系的植物;(b)提供第二馬鈴薯植物,其中所述第二馬鈴薯植物是可具有任何純合水平的二倍體或四倍體馬鈴薯品系的植物,所述第二馬鈴薯植物的品系包括具有以每株植物至少200克鮮重的克數表示的平均塊莖產量的植物;(c)使所述第一馬鈴薯植物和第二馬鈴薯植物異花授粉以提供種子,且收集所述種子從而提供雜種后代種子形式的后代;(d)使所述雜種后代種子長成雜種后代馬鈴薯植物的種群,且從以每株植物至少200克鮮重的克數表示的產量產生塊莖的所述種群的植物中進行選擇;以及(e)使所述雜種后代馬鈴薯植物自交或使所述雜種后代馬鈴薯植物與所述第一馬鈴薯植物的品系的馬鈴薯植物回交,進行1至8次自交或回交,從而提供二倍體、自交親和且基本純合的馬鈴薯育種品系,該二倍體、自交親和且基本純合的馬鈴薯育種品系具有以每株植物至少200克鮮重的克數表示的平均塊莖產量。本文所述方法中的第一馬鈴薯植物優選為二倍體、自交親和、能育且基本純合的第一馬鈴薯品系的植物,其中所述自交親和性受到s基因座(自交不親和基因座)抑制基因(s-locusinhibitorgene,sli)的顯性等位基因存在的控制,其中sli基因座抑制基因是如hosaka和hanneman(1998euphytica,99:191–197)描述的s基因座的非等位基因。當以足夠大的規模(至少超過10000株植物)且使用適當的第二馬鈴薯植物實施上述方法時,且該第二馬鈴薯植物優選為基本不純合的二倍體馬鈴薯克隆,所述第二馬鈴薯植物的品系包括具有以每株植物至少200克鮮重的克數表示的平均塊莖產量的植物,優選如在下文實例中所述的ivpaa-096-18(d2)的品系,該第二馬鈴薯植物的品系會生產出基本純合的、能育的、有活力且自交親和的育種品系,其中所述活力尤其以每株植物至少200克鮮重的克數表示的平均塊莖產量表示。提供這樣的育種品系是提供基因上純合的真正馬鈴薯種子(tps)過程中的關鍵步驟。當第一育種品系和第二育種品系均包括農藝學期望特征,具有良好的一般配合力(generalcombiningability),且優選具有通過分子標記物檢驗可確定的至少20%的相對的純合的基因座時,第一育種品系和第二育種品系隨后可用于產生tps。在根據本發明的產生二倍體、能育、自交親和且基本純合的馬鈴薯品系的所述方法的優選實施方案中,重復所述步驟a-e,其中步驟b包括提供第三(或其它的)馬鈴薯植物,其中所述第三馬鈴薯植物是具有任意純合性水平的二倍體或四倍體馬鈴薯品系的植物,其中所述第三馬鈴薯植物是額外的農藝學期望性狀的供體。優選地,所述性狀選自由昆蟲耐性、線蟲抗性、疾病抗性(包括但并不限于對粉痂病、絲核菌、銀屑病、晚疫病菌、鏈霉菌導致的瘡痂病的抗性)、除草劑耐性、耐寒性、耐旱性、耐濕性、耐干腐和濕腐性、耐鹽性及對低溫糖化的抗性,或本文所述的任何其他農藝學有益的性狀或期望的特征所組成的組中。上述第二馬鈴薯植物或第三馬鈴薯植物可適當地選自下列實例3的表6中所列的植物d1~d21的二倍體馬鈴薯克隆。技術人員應理解也可使用其它二倍體馬鈴薯克隆。非常優選的是使用馬鈴薯克隆ivpaa-096-18(d2),其可從荷蘭瓦赫寧根大學的植物育種實驗室(laboratoryofplantbreeding(ivp),wageningenuniversity,droevendaalsesteeg1,6708pbwageningen,thenetherlands)的irronaldb.c.hutten博士處獲取。四倍體栽培品種的適當(商品化)來源是歐洲栽培馬鈴薯數據庫(europeancultivatedpotatodatabase,ecpd)(http://www.europotato.org/menu.php),和/或專家組植物育種的馬鈴薯譜系數據庫(與荷蘭瓦赫寧根大學植物育種實驗室(laboratoryofplantbreedingofwageningenur,thenetherlands)之間的合作),以及荷蘭瓦赫寧根國際植物研究所的生物多樣性及育種業務單位(pri)(http://www.plantbreeding.wur.nl/potatopedigree)。在所述方法的另一優選實施方式中,所述第一馬鈴薯植物是馬鈴薯品系ncimb41663、ncimb41664、ncimb41665或ncimb41765的植物,所述品系的代表性種子保藏于蘇格蘭阿伯丁的英國食品工業與海洋細菌菌種保藏中心(ncimb)。在所述方法的另一優選實施方式中,所述第二馬鈴薯植物是二倍體、能育、自交親和且基本純合的第二馬鈴薯品系的植物。在所述方法的再一優選實施方式中,所述第二馬鈴薯植物是二倍體、能育且自交親和的第二馬鈴薯品系的植物。用于產生二倍體馬鈴薯育種品系的方法可進一步包括:通過互交、自交及回交增強二倍體馬鈴薯育種品系的育種價值,并針對農藝學期望性狀進行連續選擇的步驟。進行試驗雜交以評價一般配合力(互交能力)。另一方面,本發明提供了一種通過上述本發明的方法獲得的二倍體馬鈴薯育種品系,更優選為良種品系。另一方面,本發明提供了一種雜種馬鈴薯種子,所述雜種馬鈴薯種子是由上述根據本發明的第一二倍體馬鈴薯育種品系的植物和第二二倍體馬鈴薯育種品系的植物雜交獲得,其中通過分子標記物檢驗確定所述第一二倍體馬鈴薯育種品系和所述第二二倍體馬鈴薯育種品系的植物包含至少20%的相對的純合的基因座。另一方面,本發明提供了一種用于產生一致的雜種馬鈴薯種子的方法,包括:(a)提供第一馬鈴薯植物,其中所述第一馬鈴薯植物是二倍體、能育、自交親和且基本純合的第一馬鈴薯品系的植物,(b)提供第二馬鈴薯植物,其中所述第二馬鈴薯植物是二倍體、能育、自交親和且基本純合的第二馬鈴薯品系的植物,其中通過分子標記物檢驗確定所述第一植物和第二植物包含至少20%的相對的純合的基因座,(c)使所述第一馬鈴薯植物和第二馬鈴薯植物進行異花授粉,且允許所述親本植物中的一株產生具有種子的漿果,并從所述漿果中收集所述種子以提供雜種后代種子形式的后代。在用于產生雜種種子的方法的優選實施方式中,所述第一馬鈴薯植物和第二馬鈴薯植物源自包括具有以每株植物至少200克鮮重的克數表示的平均塊莖產量的植物的馬鈴薯品系,優選其中所述第一馬鈴薯植物和第二馬鈴薯植物是塊莖產量為每株植物至少200克鮮重的植物。在產生雜種種子的方法的另一優選實施方式中,所述第一馬鈴薯植物和第二馬鈴薯植物中的至少一個包括至少一種農藝學期望的性狀,所述農藝學期望的性狀優選選自由昆蟲耐性、線蟲抗性、疾病抗性(包括但并不限于對粉痂病、絲核菌、銀屑病、晚疫病菌、鏈霉菌導致的瘡痂病的抗性)、除草劑耐性、耐寒性、耐旱性、耐濕性、耐干腐和濕腐性、耐鹽性及對低溫糖化的抗性所組成的組中。在根據本發明的用于產生雜種種子的方法的另一優選實施方式中,所述第一馬鈴薯品系是含有sli基因的、二倍體、自交親和且基本純合的后代馬鈴薯植物,優選其中所述第一馬鈴薯植物是馬鈴薯品系ncimb41663、ncimb41664、ncimb41665或ncimb41765的植物,或是所述植物的二倍體、自交親和且基本純合的后代馬鈴薯植物,所述品系的代表性種子保藏于蘇格蘭阿伯丁的ncimb。在根據本發明的用于產生雜種種子的方法的另一優選實施方式中,所述第二馬鈴薯植物是二倍體、自交親和且基本純合的第二馬鈴薯品系的植物,優選包括至少兩個農藝學期望的性狀,所述農藝學期望的性狀選自由昆蟲耐性、線蟲抗性、疾病抗性(包括但并不限于對粉痂病、絲核菌、銀屑病、晚疫病菌、鏈霉菌導致的瘡痂病的抗性)、除草劑耐性、耐寒性、耐旱性、耐濕性、耐干腐和濕腐性、耐鹽性及對低溫糖化的抗性所組成的組中。另一方面,本發明提供了一種馬鈴薯品系ncimb41663、ncimb41664、ncimb41665或ncimb41765的種子,所述品系的代表性種子保藏于蘇格蘭阿伯丁的ncimb。另一方面,本發明提供了一種通過本文所公開的用于產生一致的雜種馬鈴薯種子的方法所產生的種子。另一方面,本發明提供了一種由本發明的種子長成的雜種馬鈴薯植物或其一部分。另一方面,本發明提供了一種本發明的雜種馬鈴薯植物的花粉或胚珠,或其一部分。另一方面,本發明提供了一種來自本發明的雜種馬鈴薯植物的塊莖。另一方面,本發明提供了一種具有由本發明的種子長成的雜種馬鈴薯植物的所有生理和形態特征的馬鈴薯植物。另一方面,本發明提供了一種本發明的雜種馬鈴薯植物的組織培養物。另一方面,本發明提供了一種由本發明的組織培養物再生得到的馬鈴薯植物,其中所述再生得到的馬鈴薯植物具有由本發明的種子長成的雜種馬鈴薯植物的所有生理和形態特征。另一方面,本發明提供了一種用于產生雜種馬鈴薯種子的方法,其中所述方法包括使第一馬鈴薯植物與第二馬鈴薯植物雜交,并收獲得到的雜種馬鈴薯種子,其中所述第一親本馬鈴薯植物和/或第二親本馬鈴薯植物是根據本發明的二倍體、能育、自交親和且基本純合的植物,該植物具有以每株植物至少200克鮮重的克數表示的平均塊莖產量。另一方面,本發明提供了一種產生雜種馬鈴薯植物的方法,所述方法包括根據本發明的方法產生種子并使所述種子長成馬鈴薯植物。另一方面,本發明提供了一種通過本發明的方法產生的馬鈴薯植物,或其塊莖。另一方面,本發明提供了至少4種可互交的、純合的、二倍體、能育、自交親和且基本純合的馬鈴薯品系的方法組,所述馬鈴薯品系包括根據本發明的具有以每株植物至少200克鮮重的克數表示的平均塊莖產量的植物,其中每個所述品系與所述組中的另一個品系的不同之處在于具有根據分子基因標記物檢驗可確定的至少20%的相對的純合基因座。本發明還提供了作為本發明上述方法的實施方式的方法,包括以下步驟:(a)提供第一馬鈴薯植物,所述第一馬鈴薯植物是二倍體、自交親和且基本純合的第一馬鈴薯品系的植物,且所述第一馬鈴薯品系的植物攜帶具有要在后代中表現出來的農藝學期望性狀的等位基因的染色體,(b)提供第二馬鈴薯植物,其中所述第二馬鈴薯植物是二倍體、自交親和且基本純合的第二馬鈴薯品系的植物,且所述第二馬鈴薯品系的植物攜帶具有要在后代中表現出來與所述第一植物的性狀不同的農藝學期望性狀的等位基因的染色體,(c)使所述第一馬鈴薯植物和第二馬鈴薯植物進行異花授粉,使得所述親本植物中的一個親本植物產生具有種子的漿果,并從所述漿果中收集所述種子以提供雜種后代種子形式的、攜帶具有所述等位基因的染色體組合的后代。因此,在可選擇的實施方式中,第一馬鈴薯品系的植物可攜帶具有要在后代中表現出來的農藝學期望性狀的等位基因的染色體,在這種情況下,第二馬鈴薯品系的植物的農藝學期望性狀可不同于所述第一植物的性狀。另一方面,本發明提供了一種產生抗除草劑、抗線蟲、抗昆蟲和/或抗病的馬鈴薯植物的方法,所述方法包括根據上述的本發明方法通過使抗除草劑、抗線蟲、抗蟲和/或抗病的親本品系雜交從而產生種子,并使所述種子長成馬鈴薯植物。另一方面,本發明提供了一種通過上述的本發明方法所產生的抗除草劑,抗線蟲,抗昆蟲和/或抗病的馬鈴薯植物。另一方面,本發明提供了一種防止病毒感染在馬鈴薯的世代之間傳播的方法,包括:(a)提供第一馬鈴薯植物,其中所述第一馬鈴薯植物是第一二倍體、自交親和且基本純合的馬鈴薯品系的植物;(b)提供第二馬鈴薯植物,其中所述第二馬鈴薯植物是具有任意倍性和/或任意純合水平的馬鈴薯品系的植物,所述第二馬鈴薯植物攜帶具有要在后代中表現出來的農藝學期望性狀的等位基因的染色體;(c)使所述第一馬鈴薯植物和第二馬鈴薯植物進行異花授粉,使得所述親本植物中的一個親本植物產生具有種子的漿果,并從所述漿果中收集所述種子以提供雜種后代種子形式的攜帶具有所述等位基因的染色體的后代;(d)使所述雜種后代種子長成攜帶具有所述等位基因的染色體的雜種后代馬鈴薯植物;(e1)從所述雜種后代馬鈴薯植物收獲塊莖以提供表現出所述性狀的塊莖;(e2)使所述雜交后代馬鈴薯植物自交或使所述雜交后代馬鈴薯植物與所述第一馬鈴薯植物的品系的馬鈴薯植物回交,從而進一步提供攜帶具有所述等位基因的染色體的后代植物,其中所述后代植物在每次回交或自交步驟之后逐漸增加純合性。另一方面,本發明提供了一種二倍體、能育、自交親和且基本純合的馬鈴薯品系,所述馬鈴薯品系包括具有以每株植物至少200克鮮重的克數表示的平均塊莖產量的植物,所述植物通過以下方式獲得:(a)提供第一馬鈴薯植物,其中所述第一馬鈴薯植物是第一二倍體、能育、自交親和且基本純合的馬鈴薯品系的植物,(b)提供第二馬鈴薯植物,其中所述第二馬鈴薯植物是具有以每株植物至少200克鮮重的克數表示的平均塊莖產量的二倍體馬鈴薯品系的植物,(c)使所述第一馬鈴薯植物和第二馬鈴薯植物進行異花授粉,允許所述親本植物中的一個親本植物產生具有種子的漿果,并從所述漿果中收集所述種子以提供雜種后代種子形式的后代,(d)使所述雜種后代種子長成雜種后代馬鈴薯植物,(e)使所述雜種后代馬鈴薯植物與所述第一馬鈴薯植物的品系的馬鈴薯植物回交,且從后代植物中進行選擇,從而生成二倍體、自交親和且有活力的植物,因為該植物以每株植物至少200克鮮重的克數表示的產量產生塊莖,(f)使所述二倍體、自交親和且有活力的植物自交1至8次,同時針對(e)中所述的自交親和和活力進行選擇。另一方面,本發明提供了一種雜交馬鈴薯種子,播種時該馬鈴薯種子產生具有以每株植物至少200克鮮重的克數表示的平均塊莖產量的二倍體、能育、自交親和且基本純合的馬鈴薯植物,所述植物通過以下方式獲得:(a)提供二倍體、能育、自交親和且基本純合的第一馬鈴薯品系,所述第一馬鈴薯品系包括具有以每株植物至少200克鮮重的克數表示的平均塊莖產量的植物,(b)提供二倍體、能育、自交親和且基本上純合的第二馬鈴薯品系,所述第二馬鈴薯品系包括具有以每株植物至少200克鮮重的克數表示的平均塊莖產量的植物,其中通過分子標記物檢驗確定所述第一植物和第二植物包含至少20%的相對的純合的基因座,(c)使所述第一馬鈴薯植物和第二馬鈴薯植物進行異花授粉,允許所述親本植物中的至少一個親本植物產生具有種子的漿果,并從所述漿果中收集所述種子以提供雜種后代種子形式的后代。選擇或提供純合植物的步驟,或選擇或提供含有本文所定義的至少20%的相對基因座的植物的步驟,可以包括利用本文提供的分子基因標記物進行標記物輔助選擇。附圖說明圖1示出了利用本發明的方法和植物來開發雜種馬鈴薯栽培品種的開發方案的示意圖,其中示出了獲得商品化規模的馬鈴薯雜交種的各個步驟:0,馬鈴薯種質:起源原則上包括種植的馬鈴薯物種(solanumtuberosum)及相關的可雜交物種中的所有馬鈴薯基因型;1,自交親和的二倍體馬鈴薯:種植的馬鈴薯(solanumtuberosum)通常為自交不親和馬鈴薯。從相關物種引入自交親和性(例如參見hermsen,j.g.th.1978,euphytica27,1-11);2,純合、能育、自交親和的二倍體馬鈴薯:可通過花藥培養(jacobsen和sopory,1977,theor.appl.genet.52,119-123),通過胚珠培養或通過重復自交(phumichai&hosaka,2006,euphytica149,251-258)使單倍體加倍(uijtewaal等,1987,theor.appl.genet.73,751-78),來獲得純合二倍體,本文闡述了重復自交的實例;3,有活力的、能育、純合、自交親和的二倍體馬鈴薯:通過與其他二倍體馬鈴薯雜交、自交及回交并選擇較高活力的個體,來提高活力;4,二倍體馬鈴薯育種品系:通過與其他二倍體馬鈴薯雜交、自交及回交并選擇具有農藝學期望性狀的個體,來引入這些性狀;5,優秀的二倍體馬鈴薯育種品系:通過互交、自交及回交并連續選擇具有農藝優良性狀的個體,來提高育種價值。進行試驗性雜交以評價一般配合力;6,馬鈴薯雜交栽培品種:對良好的配合育種品系進行雜交,獲得的種子為一致的雜種真正馬鈴薯種子。圖2示出了實例中所述種群,保藏及植物的系譜的示意圖,從二倍體、基本純合且自交親和的馬鈴薯植物(“h”)與三種不同二倍體克隆(d1、d2和d3)之間的雜交開始,其中所述二倍體克隆基本上非純合且自交不親和且充當一個或多個期望農藝學性狀的供體品系。從回交品系至全自交克隆產生一系列基本純合的品系,每個品系均具有一個或多個源自供體品系的期望性狀,從而提供一組用于產生一系列雜種種子形式的雜種馬鈴薯栽培品種的優良育種品系。表示自交。如圖可看出,并如實例中詳細描述的,只有供體品系d2得到了表現如本文所述活力的自交親和植物的大量產生。有活力的植物可從其他雜交組合獲得,但頻率會很低(0.1-1%)。圖3顯示了二倍體育種克隆rh89-039-16的基因組的超密基因連鎖圖譜(ultra-densegeneticlinkagemap)。連鎖群左邊的數字是從頂部開始計算的重組事件的累積次數。每個框(bin)中的aflp(隨機擴增長度多態性)標記物的數量用根據圖例的灰色陰影表示。推定的著絲粒位置用染色體旁邊的“cen”表示(vanos等,2006genetics173(2):1075-1087)。連鎖群右邊的數字表示本文所用的對應的potsnp標記物的大體位置,其中參照下列實例3中的表5b獲取標記物的注解。附圖清楚地顯示提供信息的標記物在該克隆的馬鈴薯基因組上的分布。利用本文所用的另一克隆sh83-92-488觀察這些標記物的類似分布。圖4示出了100種標記物在20種不同二倍體馬鈴薯基因型上的標記物數據,表示二倍體馬鈴薯育種的種質(實例2)。圖5示出了對一組六個f3種群及各自的親本進行如在實例3中描述的snp標記物檢驗的結果。具體實施方式定義本文使用的術語“馬鈴薯”指基本是solanumtuberosum種類的材料,但可包括其它結出塊莖的茄屬植物(如solanumchacoense、solanumphureja、solanumandigena、solanumdemissum)的基因滲入的部分。本文使用的術語“馬鈴薯植物”指如在細胞培養或種子生長的幼苗或成熟的植物。本領域技術人員應識別當術語“馬鈴薯植物”用于本發明的上下文中時,還包括保持為本發明主題的例如馬鈴薯品種的基本區別特種的衍生品種,如該種類的單基因轉化的植物(singlegeneconvertedplant)或具有一個或多個并入其中的增值基因(如除草劑或害蟲抗性)的轉基因衍生物。回交方法可以與本發明一起使用以改善品種或將特征引入品種。如本文使用的術語“雜交”指雌株(或配子)被雄株(或配子)受精。術語“配子”指在植物中通過有絲分裂從配子體產生的且參與有性繁殖的單倍體生殖細胞(卵子或精子),在有性繁殖過程中異性的兩個配子融合以形成雙倍體合子。該術語通過包括花粉(包括精子細胞)和胚珠(包括卵)。因此,“雜交”通常指一個個體的胚珠經來自另一個體的花粉受精,然而“自交”指一個個體的胚珠經來自相同個體的花粉受精。雜交廣泛用于植物育種中,且導致雜交的兩株植物之間的基因組信息的混合,一個染色體來自母親而另一染色體來自父親。這將導致基因遺傳性狀的重新組合。雜交的后代一般被定名為“f1”。如果f1不是一致的(分離的),則一般被定名為“f1種群”。純合植物的“自交”一般將導致基因一致的植物,因為沒有基因變異。f1的“雜交”將導致在f1中具有雜合基因座的所有性狀分離的后代。這樣的后代被定名為“f2”或“f2種群”。當在實現基因組區或段的基因滲入的環境中提到“雜交”時,技術人員應理解由于在親本品系中的配子產生中發生的交換事件,為了實現一株植物的僅染色體的一部分基因滲入另一植物的染色體中,需要兩個親本品系的基因組的隨機部分在雜交過程中均進行重組。因此,兩個親本的基因組必須通過雜交組合在一個細胞中,之后,來自所述細胞的配子的產生及它們在受精中的融合將導致基因滲入事件發生。如本文所使用的術語“可互交的”指在親本植物之間雜交之后產生后代植物的能力。如本文所使用的術語“基因滲入中(introgressing)”、“基因滲入”和“基因滲入的”指天然和人工的過程,借此單個基因或整條染色體通過使個體、物種、品種或栽培品種進行雜交而從一個個體、物種、品種或栽培品種被移入另一個體、物種、品種或栽培品種的基因組中。在植物育種中,該過程一般包括自交或回交至回歸親本以提供基本具有除了基因滲入的機或性狀之外的回歸親本的特征的漸增的純合植物。術語“基因滲入”指基因滲入事件的結果。術語“回交品種(backcross)”指“回交”過程的結果,其中,從在兩個親本品系之間的雜交所得到的植物(重復)與它的親本品系之一雜交,其中在回交中使用的親本品系指為回歸親本。重復回交導致給方親本(donorparent)的基因組片段被回歸哪些基因組片段所取代。雜交后代被定名位“bcx”或“bcx種群”,其中“x”表示回交的次數。如本文所使用的術語“回交”指雜交后代后退與回歸親本的重復雜交。把為期望特征貢獻基因的親本馬鈴薯植物叫做非回歸親本或給方親本。該術語指非回歸親本在回交方案中使用一次且因此不會重現的事實。來自非回歸親本的單個基因或多個基因被轉移至其中的親本馬鈴薯植物已知為回歸親本,因為它在回交方案中被使用幾個循環。在典型的回交方案中,感興趣的原始品種(回歸親本)與攜帶要被轉移的感興趣的單個基因的第二品種(非回歸親本)雜交。然后,從該雜交所得到的后代再次與回歸親本雜交,且重復該過程直至獲得馬鈴薯植物,其中除了從非回歸親本轉移的單個基因或有限數量的基因之外,在被轉變的植物中基本恢復了回歸親本的所有期望的表型和生理特征。選擇適合的回歸親本對于成功的回交程序是重要的步驟。回交方案的目標是改變或取代原始品種中的單個性狀或特征。為了實現這一目的,通過來自非回歸親本的期望基因對回歸品種的單個基因進行修飾、取代或補充,同時基本保持所有剩余的期望基因,且因此保持原始品種的期望的生理和形態構成。具體的非回歸親本的選擇將依賴于回交的目的。其中一個主要的目的是向回歸親本中加入一些商業上期望的、農藝學上重要的性狀。精確的回交方案將依賴于要改變或加入的特征或性狀以確定適合的測試方案。雖然當時被轉移的特征是顯性等位基因時簡化了回交方法,但還可能轉移隱性等位基因。在這種情況下,可能需要引入后代的測試以確定期望的特征是否已經被成功轉移。優選地,通過診斷的分子標記物監控這樣的基因。同樣地,可以使用本領域技術人員已知的各種已建立的重組方法中的任何方法將轉基因引入植物中。已經鑒定出很多單個基因的性狀,這些性狀在新型品種的開發中不是定期選擇的,但通過回交和基因工程技術是可以改善的。單個基因性狀可以或可以不是轉基因的,例如這些性狀包括但并不限于:除草劑抗性,對細菌性疾病、真菌性疾病或病毒性疾病的抗性,昆蟲抗性,淀粉和其它碳水化合物濃度的均勻性或淀粉和其它碳水化合物濃度的增加,增強的營養品質,降低塊莖易于碰傷的傾向,及降低淀粉轉變為糖的比率。術語“自交”指自受精的過程,其中,個體被自己的花粉授粉或受精。重復自交最終導致純合的后代。如本文所使用的術語“品系”指來源于單次雜交、回交或自交的植物的種群。個體后代植物不會必然與另一個相同。個體后代植物由于變異可能沒有活力、不能育的或不可自交。但是,預見到可以在品系中容易地鑒定出有活力的、能育的或可自交的適合的植物,且將該適合的植物用于額外的育種目的。如在本文中所使用的,術語“等位基因”意味著基因的一個或多個可選擇形式中的任意一個,所有這些等位基因均涉及至少一個性狀或特征。在雙倍體細胞或有機體中,兩個拷貝的基因占據一對純合染色體上的對應的基因座。每一拷貝可以是有區別的等位基因。本文中的“基因”被定義為遺傳單位(常常由dna序列表示),該遺傳單位占據染色體上的特定定位,且包含用于對植物中的潛在表型特征或性狀作貢獻的基因指令。本文中的“基因座”被定義為給定基因在給定植物物種的染色體上占據的位置。如本文所使用的,術語“純合的”意味著當相同的等位基因存在于同源染色體上的對應基因座上時存在的基因條件。術語“基本純合的”指當測試100、優選1000、更優選至少10000個基因座時,具有至少25%,優選至少50%,更優選至少75%,還更優選至少76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99或100%的純合性的水平。技術人員應理解植物的平均(level)純合性由植物的整個基因組所顯示出的純合性水平來定義,且如通過基因座的隨機選擇所獲得的(但該測試可能依賴于所使用的標記物)測試100、優選1000、更優選至少10000基因座反映出整個植物基因組的純合性水平。純合性水平是種群的平均值,且優選指其中親本不同的那些基因座。如在本文中所使用的,術語“雜合性”意味著當不同的等位基因存在于同源染色體上的對應基因座上時存在的基因條件。術語“重組(recombination)”或“重組(recombine)”指在有絲分裂的過程中兩個同源染色體之間的信息交換。在“重組”的植物中,最初呈現在染色體內的具體定位上的dna(例如聯接至一基因/基因座)與來自另一植物(即親本中的母性或反之亦然)的dna進行交換。為了僅交換所需的材料且在染色體上盡可能多的維持有價值的原始信息,通常需要兩次側面交叉(flankingcrossover)或重組事件。在雙重重組(doublerecombination)中,該交換已經發生在基因/基因座的兩側。發現這樣的雙重重組的方式是篩選f2-植物的種群。該種群已經是大量的,因為僅以有限的頻率出現雙重重組。可選擇地,在基因單位內的雙重重組可以是連續回交的結果。雙重重組的頻率是單次重組的頻率的產物。(例如在10cm區域中的重組可以發現為10%的頻率,雙重重組發現為10%x10%=1%的頻率)。如在本文中所使用的,術語“子代(progeny)”意味著基因子孫或后代。如在本文中所使用的,術語“種群”意味著分享共同基因起源的植物的基因異種集合。“重組事件”指有絲分裂或減數分裂的交叉事件,包括gmo事件。如在本文中所使用的,術語“雜種”意味著在兩個基因上不相似的個體之間雜交的任何后代,更優選地該術語指兩個(優秀或近交的)將不真正從種子繁殖至親本的育種品系之間的雜交。如在本文中所使用的,術語“分離”指成對的等位基因在減數分裂過程中的分開,以便每對等位基因的成員出現在不同的配子中。該術語包括提及該基因現象的結果,其中,在至少一個親本對一個等位基因來說是雜合的雜交中的后代種群相對于所述基因的表型性狀是不一致的。如在本文中所使用的,術語“育種品系”指具有商業上有價值或農藝學上期望的特征的栽培的馬鈴薯的品系,與野生型品種或作物地方品種形成對照。該術語包括體積優秀的育種品系或優秀的品系,該品系表現出基本純合、通常為近交的用于產生商品化f1雜種植物品系。通過育種及對包括眾多農藝學上期望性狀的優良農藝性能的選擇獲得優秀的育種品系。優秀植物是來自良種品系的任何植物。優良農藝性能指如本文所定義的農藝學上期望的性狀的期望組合,其中期望與非優秀育種品系相比,大部分、優選所有的農藝學上期望性狀在優秀育種品系中均得到改善。優秀育種品系基本是純合的,且優選是近交品系。如在本文中所使用的,術語“良種品系”指從育種和對優良農藝性能進行選擇所得到的任何品系。良種品系優選是具有眾多,優選具有至少3、4、5、6或更多本文所定義的期望農藝性狀(的基因)的品系。術語“栽培品種”和“品種”在本文可互換使用,且指示具有通過為了商業化的目的經農民和栽培者進行育種、雜交和選擇而謹慎開發出的植物,以產生農業產物用于自己消費或用于商用(新鮮消費、處理、喂養等)。術語“育種種質”指示具有除了“野生型”狀態植物的生物狀態的植物,該“野生型”狀態指植物或近似物(accession)的原始未經栽培或天然的狀態。術語“育種種質”包括,但并不限于:半天然、半野生型、雜草類的、傳統的栽培品種、作物地方品種、育種材料、研究材料、培植者的品系、人造種群、雜種、創造者原種(founderstock)/基礎種群、近交品系(雜種栽培品種的親本)、分離的種群、突變體/基因原種、高級市場和先進/改善的栽培品種。栽培品種的實例包括栽培的品種,如bintje、russetburbank、eigenheimer和nicola。如在本文中所使用的,術語“純種的”、“純種近交”或“近交”可以互換,且指通過重復自交和-或回交獲得的基本純合的植物或植物品系。如在本文中所使用的,術語“分子基因標記物”或短的“標記物”指在用于使在核酸序列特征中的差異可視化的方法中使用的指示物。這樣的指示物的實例是限制性片段長度多態性(rflp)標記物、擴增片段長度多態性(aflp)標記物、單核苷酸多態性(snp)、插入/缺失(indel)突變、微衛星標記物(ssr)、序列特異擴增區域(scar)、剪切擴增多態性序列(caps)標記物或同功酶標記物或本文所描述的限定特定基因和染色體定位的標記物的組合。如在本文中所使用的,術語“植物部分”指示馬鈴薯植物的一部分,包括細胞器、單個細胞和細胞組織,如植物中完整的植物細胞、細胞團和組織培養物,從該部分中可以再生出馬鈴薯植物。植物部分的實例包括,但并不限于:來自花粉、胚珠、葉子、胚、根、根尖、塊莖、花藥、花、漿果、莖桿(stemshoot)和種子的單個細胞和組織;以及花粉、胚珠、葉子、胚、根、根尖、花藥、花、漿果、莖、芽(shoot)、穗木、根莖、種子、塊莖、原生質體、愈傷組織等類似物。除非特別聲明,術語“種子”在本說明書中均指從中長出新型植物的物體(或一些植物中的籽粒),包含通常與一些儲存的食物在一起的、包在種皮覆蓋(seedcoatcovering)中的小的胚性植物。該種子(被稱為植物的或“真正”的種子)是裸子植物和被子植物的成熟胚珠的產物,在受精且在母性植物內生長一段時間后出現。如在本文中所使用的,術語“活力”和“有活力的”指植物的地上組織或地下組織的相對量,該相對量可以是彼此或多或少地不互相依賴。地上組織的相對量可以表示為按照以等級分類評定的植物高度、莖的數量、葉子的數量,和/或按照鮮重或干物質的枝葉的量的可視化觀察結果。該性狀在植物生長周期的第一階段的過程中具有關聯,該植物生長周期在五月至八月的季節中歐洲西北部普遍為在正常生長條件下具有100天的平均種植時間(75~130天的范圍)。在這些條件下,本發明的植物獲得以鮮重或干重(克)表示的枝葉和芽(地上植物)的重量,該重量為(基本雜合的)二倍體育種克隆rh89-039-16的至少50%、更優選為至少60%、70%、80%、90%或更多,反過來該重量具有基于九年測試的四倍體馬鈴薯品種的塊莖產量的68%(44~86%的范圍)。克隆rh89-039-16可用于irronaldb.c.hutten博士(植物育種實驗室,瓦格寧根大學,droevendaalsesteeg1,6708pb瓦赫寧恩,荷蘭)的目的。可選擇地或額外地,術語活力可以與特定最小塊莖產量相連使用。如在本文所使用的,術語“塊莖產量”將一般指在五月至八月的季節中在歐洲西北部流行的具有正常生長條件的100天栽培時間段的條件下獲得的以鮮重(克)表示的塊莖產量,該塊莖產量在根據本發明的植物收獲時為(基本雜合的)二倍體育種克隆rh89-039-16的至少30~50%、更優選為至少60%、70%、80%、90%或更多,反過來該塊莖產量具有基于九年大田測試的四倍體馬鈴薯品種的塊莖產量的68%(44~86%的范圍)。如上所示,克隆rh89-039-16可用于irronaldb.c.hutten博士(植物育種實驗室,laboratoryofplantbreeding,瓦格寧根大學,droevendaalsesteeg1,6708pb瓦赫寧恩,荷蘭)的目的。除非有特別表示,與從塊莖生長的植物形成的塊莖形成對照,塊莖產量是基于從種用植物形成的塊莖,該產量可以比種用產量高50~200%。可選擇地或額外地,術語活力可以指以鮮重或干重(克)表示的塊莖產量,該塊莖產量在收獲時是二倍體育種克隆rh89-039-16的至少30~50%、更優選為至少60%、70%、80%、90%或更多。這意味著在粘質土中在正常歐洲西北部夏天的條件下,(在本發明的植物中)以鮮重(克)表示的塊莖產量在每株從植物種子生長的植物中可以達到至少200克、更優選至少300克、400克或500克,且當所述植物從塊莖種植且在粘質土中在正常歐洲西北部夏天的條件下生長時,甚至達到每株植物中為400克、500克、600克、700克、800克或更多,如900克或1000克的新鮮塊莖重量或更多。術語“包括植物的品系”中所述植物指為具有特定特征(如塊莖產量),且術語“包括植物的品系”應優選理解為指基本由具有所述性狀、但允許一些生物變異的植物所組成的品系。如在本文中所使用的,術語“二倍體”指其中每一營養細胞(vegetativecell)均包含兩組染色體(2x=2n,其中n是染色體的數量)的植物。染色體中的一組來自各自的親本。如在本文中所使用的,術語“四倍體”指其中每一營養細胞均包含四組染色體(2x=4n)的植物。如在本文中所使用的,術語“線蟲抗性”指已經基因滲入功能抗性基因以預防至少一種線蟲種群或隔離種群的增殖的植物。如在本文中所使用的,術語“疾病抗性”指示出使發病葉面或塊莖體積的量,或昆蟲或病原微生物(包括但并不限于鏈霉菌屬(streptomycesspp)、絲核菌(rhizoctonia)、銀屑病(silverscurf)和晚疫病菌(phytophthorainfestans))增殖的量降低高于50%的能力。如在本文中所使用的,術語“除草劑耐性”指比在應用規定劑量的除草劑所通常導致的損害示出低于50%的枝葉損害的植物。如在本文中所使用的,術語“耐寒性”指示出霜凍害的植物葉面的平均量,在耐寒植物中的損害低于相對冷敏感的參照品種(如caribe和/或kennebec)在零下3℃的溫度下所觀察到的平均損害。如在本文中所使用的,術語“耐旱性”指與相對干旱敏感的參照品種(如caribe和/或carlton)相比,在限制性水條件下示出低于平均損害的植物。如在本文中所使用的,術語“耐澇性”指在洪水泛濫時受影響的塊莖(厭氧降解)的比例相對于示出的平均損害的植物低50%。如在本文中所使用的,術語“耐濕腐性”指對歐文氏菌屬(erwinia)(目前被叫做果膠桿菌屬(pectobacteriumspp.))具有最低水平的抗性的植物。如在本文中所使用的,術語“耐干腐性”指對鐮刀菌屬(fusarium)具有最低水平抗性的植物。如在本文中所使用的,術語“耐鹽性”指在鹽性條件下示出比平均損害較低的損害的植物。如在本文中所使用的,術語“生長率”指每一時間單位植物生物量的增加。如在本文中所使用的,術語“塊莖發育缺陷”(例如畸形或受損的塊莖)指在植物發育和/或塊莖收獲的過程中示出塊莖的看得見及內部的變形。如在本文中所使用的,術語“塊莖產量”指通常以鮮重(克)表示的植物塊莖或植物種群的總重量。如在本文中所使用的,術語“塊莖大小”指塊莖的絕對寬度、高度和長度。有益的塊莖大小包括每個塊莖大約80~160克的植物平均塊莖大小。商業可用的植物的平均塊莖數是每株植物大約8~12個塊莖。如在本文中所使用的,術語“塊莖皮的顏色”指由于塊莖皮組織中花色素苷的積累而導致塊莖皮在收獲后的顏色。如在本文中所使用的,術語“芽眼深度(eyedepth)”指塊莖的皮表面和芽原基之間的相對距離。如在本文中所使用的,術語“塊莖形狀”指:長度/寬度的比率,以指示從圓形、橢圓形至長塊莖形狀的持續變異;以及高度/寬度的比率,以指示塊莖從圓柱形至扁平量的持續變異。如在本文中所使用的,術語“塊莖果肉的顏色”指由于缺失或存在類胡蘿卜素化合物分別引起白色或黃色果肉顏色,以及缺失或存在花色素苷化合物分別引起果肉顏色的紅色、藍色、紫色陰影所造成無論顏色是否以部分或全部的模式存在的塊莖果肉在收獲后的內部的顏色。如在本文中所使用的,術語“塊莖味道”指通過吃蒸煮的馬鈴薯塊莖消費者的評價。如在本文中所使用的,術語“塊莖保存期限”和“塊莖儲存能力”是同義的,且指塊莖在儲存時外貌沒有變化。如在本文中所使用的,術語“塊莖休眠期”指在塊莖播種和發芽之間的時間段,同時在正常條件下播種。如在本文中所使用的,術語“塊莖脫水抗性”指示出比在濕度低的條件(見保存期限)下塊莖的平均脫水低的塊莖脫水的植物。如在本文中所使用的,術語“塊莖淀粉含量”指淀粉重量比塊莖總鮮重。如在本文中所使用的,術語“塊莖干物質含量”指馬鈴薯的干組分的重量除以總鮮重。如在本文中所使用的,術語“塊莖蒸煮品質”指塊莖在蒸煮后從堅硬至粉狀的持續變異。如在本文中所使用的,術語“塊莖油炸品質”指消費者對塊莖在油炸后的評價。如在本文中所使用的,術語“塊莖制片的品質”指消費者對塊莖被制片后的評價。如在本文中所使用的,術語“塊莖的一致性”指塊莖性狀的標準偏差,從而低標準偏差指高度不一致且反之亦然。如在本文中所使用的,術語“對低溫糖化的抗性”指當塊莖塊莖儲存在低于8℃的溫度下,對產生葡萄糖和/或果糖的抗性。如在本文中所使用的,術語“四倍體”指其中每一營養細胞均包含四套染色體(4n)的植物。術語“能育的”和“能育性”寬泛地指繁殖能力,即懷孕(conceive)和產生后代、優選能育后代的能力。因此,術語能育性指可以雜交以產生能育后代的個體,因為當授粉后,花粉產生包含種子的漿果,種子播種后會產生自身攜帶能育花等的植物。術語“能育的后代”或“能育的種子”在本文被定義為能夠長成產生花的馬鈴薯植物的種子,其中,花是雌性和雄性能育的。因此,該術語優選指植物或種子,該植物或種子當長成植物時能夠借助能育胚珠和能育花粉(即雄性和雌性花均是能育的)的存在產生作為雄性和雌性親本的后代。術語“能育的植物”在本文被定義為攜帶漿果的能夠產生種子的植物。優選地,每一所述漿果攜帶至少5、更優選至少15~20、甚至更優選至少50個能育的種子,仍然更優選攜帶50至500個之間的種子。可選擇地,植物被稱為能育的,當每一漿果產生的種子數量是在二倍體育種克隆rh89-039-16的漿果中發現的數量的至少10%、更優選至少30%、40%、50%、60%、70%、80%、90%、100%、150%、200%或更高。該術語還(或可選擇地)指具有多于25%、優選多于90%的能育植物的植物品系。術語“自交親和的”指使種子在漿果中發育的能力,是自授粉、自受精的結果且產生能育后代。術語“能夠回交/自交/雜交且得到(自交)能育的后代和自交親和的后代”指作為回交、自交或雜交的結果產生一個或多個具有種子的漿果的能力,其中在所述一個或多個漿果中超過60%、優選超過70%、80%、85%、90%或更多的種子能夠發育成(自交)能育的和自交親和的植物。對于本文中的其它術語,參考allard,r.w.的《植物育種原理(principlesofplantbreeding)第二版》(威利紐約(wileynewyork),1999),且特別參考里面的術語表。優選實施方式的描述本發明人發現可以成功產生用于馬鈴薯育種的優選育種品系。兩個優秀育種品系的雜交提供了f1雜種種子,這樣的f1雜種種子當長成植物時得到具有優良的農藝學性能的植物。更重要地,所有種子從基因方面均是基本相同的,這意味著除了偶然交換或突變事件,所有種子均是一致的。本發明人已經發現能夠供應優秀的馬鈴薯育種品系的方法,當相互雜交時該方法提供了一致的f1雜種馬鈴薯種子,該f1雜種馬鈴薯種子當長成植物時得到具有優良農藝學性能的植物。優秀的馬鈴薯育種品系可具有互補的性狀,且作為它們純合性的結果,從f1雜種種子長成的f1植物是一致的,但當這些f1植物隨后自授粉或異花授粉,所得到的f2植物將因不同性狀而被分離。因此,與從現有技術的傳統途徑產生的以及從漿果中收獲的目前可用的真正馬鈴薯種子(tps)形成對照,其中在該漿果中每一個種子從基因方面與其它種子都是不同的,從馬鈴薯品系和根據本發明各方面的優秀馬鈴薯育種品系的雜交所獲得的種子從基因方面基本是相同的。現有技術的tps品種來源于選擇的但雜合的親本之間的雜交,且由此期望到植物和塊莖中的變異性。本文中提出的雜種馬鈴薯育種與tps的傳統生產的不同在于,現在的tps是兩個純育種品系雜交的結果,產生一致的栽培品種,而現有技術的tps品種是兩個遺傳方面非純合且常常是四倍體的育種品系之間開放授粉或雜交的結果,得到不一致的栽培品種。現在提供的植物不僅在它們組合了迄今為止從來沒有在單個馬鈴薯品系中一起獲得的眾多性質方面是新穎的,而且它們還實現了新穎且非常有利的育種方法。因此,本發明的植物是產生新型馬鈴薯栽培品種的中間物,該栽培品種可以通過本文所公開的方法產生。有利的育種方法實現了消除本文上面提到的傳統馬鈴薯育種的許多缺點,包括消除克隆的耗時選擇以得到新型馬鈴薯品種,以及提供一致的雜種馬鈴薯種子的可能性。此外,新型植物和方法還實現了消除與克隆增殖和產生相關的缺點,包括消除高轉運費用及害蟲和疾病的克隆持久性的可能性。新型馬鈴薯植物親本的育種品系本發明提供了基本純合、二倍體、有活力的、能育的且自交親和的馬鈴薯植物。優選地,本發明的植物已經在它們的基因組內固定了編碼給予植物農藝學期望的性狀的基因的核酸序列。這樣的核酸序列可以是來源于具有農藝學期望的性狀的馬鈴薯種質、優選來自二倍體種質的基因滲入或轉基因。基本純合性在育種中,植物通過有性繁殖,通常通過具有期望性狀的所選擇親本的雜交進行增殖,以產生改善的子代。子代從兩個親本中遺傳得到期望性狀和不期望性狀的基因。從每一世代(將作為下一世代的親本)中重復選擇有價值的個體將保存期望的特征,且消除不期望的特征。植物育種中的最終親本是純種的高度純合的品系,該最終親本在與其它純種雜交中使用時得到在單一世代內及在從相同親本的其它雜交中獲得的連續世代之間表型和基因型均相同的后代。可以通過示出基因一致性或純合性的程度增加的每一連續世代自交或近交(接近的親緣植物的雜交,如兄弟植物對姐妹植物(brothertosisterplant)或后代對親本(回交))幾代后獲得純種或純的近交品種。作為育種親本的純合近交的使用得到在單一世代內及在這些親本的其它雜交中獲得的連續世代之間表型和基因型均相同的后代。因此,純的近交品種可重復使用以產生相同的后代植物。在馬鈴薯中,自交和近交導致嚴重的近交衰退或有害隱性基因的表達,該表達通過活力喪失和能育性喪失而顯現出來。因此,迄今已經不可能在馬鈴薯中產生純合的純的近交品種,因為直至本發明,不能在具有足夠活力的馬鈴薯中獲得高水平純合性。現在,本發明第一次提供了具有高于25%且優選高于75%的純合性水平,且組合有能育性、自交親和性和植物活力的植物。二倍體本發明的植物是二倍體植物,與為四倍體的商品化馬鈴薯品種形成對照。應注意,在本發明的植物中,明確設想了較高倍性水平,因為在育種品系的發育過程中初步使用了二倍體階段。一旦純種已經建立,則品系可以變成四倍體。因此,本發明的植物還可以是四倍體。四倍體植物可以從四倍體細胞或組織培養物獲得,反過來該四倍體細胞或組織培養物可以通過本身已知的方法使二倍體細胞或組織培養物的基因組加倍而產生,例如通過使用barandalla等.(barandalla等.potatoresearch(2006)49:143-154)描述的黃草消。四倍體植物可以具有提供較高產量的優點。有活力的雖然商品化四倍體馬鈴薯品種是能育的、自交親和的且是有活力的,因為它們生長良好,不產生變形的植物或漿果,但與具有高塊莖產量的健康植物相比,它們的自交子代示出嚴重的近交衰退,即得到具有變形塊莖的生長衰弱和低產量的植物。現有技術的二倍體和基本純合的品系(如在本文引用的phumichai等所報道的那些品系)也不僅生長非常不良,且顯示為非常弱小的具有不良的結實率(seedset)的植物。事實上,近交得到的且在等位基因上達到至少25%或50%純合性的二倍體馬鈴薯植物總是弱的生長物。本發明的植物不是弱勢和脆弱的,且雖然它們不是必然與現今商品化四倍體品種具有相同的活力,但它們沒有示出近交衰退。因此,在本發明的上下文中,它們是有活力的。塊莖形成例示了本發明的植物的活力。雖然所有商品化(四倍體和種子塊莖來源的)馬鈴薯品種產生大的塊莖,或至少具有足夠大小的塊莖,但現有技術的二倍體品系和基本純合的植物僅產生小的塊莖,該小的塊莖例如在生產薯片或炸薯條中是沒有用的。現有技術的二倍體品系和疾病純合的植物還產生非常少的塊莖,這樣每株植物的塊莖產量比雜合的二倍體植物和商品化四倍體馬鈴薯品種的塊莖產量低至少十倍。相反,本發明的植物顯示出有益的塊莖形成特征,包括有益大小外形(sizeprofile)的塊莖和塊莖產量,其塊莖產量可以是在本文中描述的實例中用作供體(d1、d2和/或d3)的二倍體雜合植物的至少50%。如本文所發現的,二倍體f1的塊莖產量平均為每株植物為443克,具有高達600克和634克的值。與f1:“ivp08-06”回交平均出產155克和209克;而與f1:“ivp08-07”回交平均出產209克和259克。“偽f2”每株植物平均出產172克和332克,且偶爾產生高于二倍體f1的產量。f2每株植物平均出產69克和252克。幾株植物與二倍體f1的產量值部分重疊。能育的本發明的植物雖然基本是純合的,但還是能育的。這意味著它們可以在育種程序中使用,因為它們的后代也將是能育的。自交親和的本發明的植物是自交親和的。引入該特征以允許植物的自交和純合品系的產生。自交親和性的優選基礎是來源于顯性s基因座的抑制基因(sli)(hosaka和hanneman,1998,euphytica,99:191–197)。本發明的基本純合的馬鈴薯植物具有如下優點:它們可以與其它基本純合的馬鈴薯植物雜交,從而所述雜交的雜種后代從遺傳方面基本是一致的,且將不會分離。在優選實施方式中,本發明提供了基本純合的、二倍體、有活力的、能育的且自交親和的具有農藝學期望性狀的馬鈴薯植物。本發明的新型馬鈴薯植物能夠:與基本純合的回歸親本回交,且得到(自交)能育的和自交親和的后代;自交(1個后代個體的自受精),且得到(自交)能育的和自交親和的后代;互相雜交(在兩個后代個體之間受精),且得到(自交)能育的和自交親和的后代。根據本發明的植物包括具有培植者編號agvd1的品系,所述品系的代表種子已經于2009年10月23日保藏在蘇格蘭阿伯丁郡的ncimb中,登錄號為ncimb41663,且培植者編號為agvd1;該品系代表通過親本ivp07-1001/4的植物與親本d1的植物雜交(見實例1),且使得到的f1后代植物自交所獲得的f2,其中親本ivp07-1001/4的植物為基本純合的、二倍體、能育的且自交可親和的但沒有活力的馬鈴薯品系,親本d1的植物為基本不是純合的,且不是自交親和的優秀的二倍體育種克隆,但親本d1的植物具有黃色果肉顏色的期望農藝學性狀且顯示出良好的蒸煮性質(因為果肉在蒸煮后不變成灰色)。估計agvd1大約50%純合,意味著致使親本不同的基因座的大約50%是純合的。根據本發明的植物還包括具有培植者編號agvd2的品系,所述品系的代表性種子已經于2009年10月23日保藏在蘇格蘭阿伯丁郡的ncimb中,登錄號為ncimb41664,且培植者編號為agvd2;該品系代表通過如上所示的親本ivp07-1001/4植物與如上所示的親本d1植物雜交,且使個體f1后代植物互相雜交所獲得的(f1xf1)品系(或偽f2),該(f1xf1)品系共享ivp07-1001/4親本但具有不同的d1親本的個體。估計agvd2對于致使親本不同的基因座大約25%是純合的。根據本發明的植物還包括具有培植者編號agvd3的品系,所述品系的代表性種子已經于2009年10月23日保藏在蘇格蘭阿伯丁郡的ncimb中,登錄號為ncimb41665,且培植者編號為agvd3;該品系代表通過如上所示的親本ivp07-1001/4的植物與如上所示的親本d1的植物雜交,且使得到的f1后代植物與ivp07-1001/4親本回交所獲得的bc1種群。估計agvd3對于致使親本不同的基因座大約75%是純合的。根據本發明的植物還包括具有培植者編號agvd17的品系,所述品系的代表性種子已經于2010年10月5日保藏在蘇格蘭阿伯丁郡的ncimb中,登錄號為ncimb41765,且培植者編號為agvd17;該品系代表通過如上所示的親本ivp07-1001/4的植物與如上所示的親本d2的植物雜交,且使得到的f1后代植物與ivp07-1001/4親本回交,然后再進行兩步自交所獲得的f3種群。估計agvd3對于致使親本不同的基因座大約80%是純合的。本發明的植物可以非常適用于用作回歸親本,其中額外的農藝學期望性狀是通過雜交基因滲入的,或通過其它如轉基因的方式引入的。一旦一些農藝學期望性狀已經被堆疊進入根據本發明的基本純合的、有活力的、二倍體、能育的且自交親和的植物中,則該植物可以被用作用于產生商品化f1雜種馬鈴薯種子的優秀育種品系。同樣地,可以提供第二優秀育種品系用作相對性別的親本,這樣兩種植物均可雜交以產生新型馬鈴薯品種的商品化f1種子。一致的雜種種子的育種依賴于一些技術條件。實現一致的雜種種子的先決條件是:i)可以在基本純合的、二倍體、能育的且自交親和的植物與作為農藝學期望性狀的供體的植物之間進行初始基因滲入的雜交;ii)可以在該初始雜交的后代與基本純合的、二倍體、能育的且自交親和的植物之間進行回交;以及,iii)其后代可以自交。馬鈴薯品系agvd1、agvd2和agvd3顯示出這些特征,且這些品系的個體植物可以在額外的基因滲入的雜交、回交和自交中使用以得到優秀育種品系,該優秀育種品系適用作用于可以長成新型馬鈴薯品種的雜種馬鈴薯植物的商品化f1雜種種子的親本。根據本發明的高度優選植物是品系agvd2的植物或其具有品系agvd2植物的所有生理和形態特征的后代。作為本發明實施方式的實例保藏的另一品系是agvd17(ncimb41765,保藏于2010年10月5日)。agvd17的優點是它是高度純合的品系(>80%的純合性),且將有活力和能育性與每株植物平均300克(鮮重)的塊莖產量相結合。相關種群的個體植物甚至示出每株植物500克的塊莖產量。這些數據由幼苗種植的大田生長的植物產生。從克隆種植的馬鈴薯植物通常產出比從幼苗種植的植物高50%~100%的塊莖重量。因此,預計在正常歐洲西北部夏天的條件下,如本文所述的植物經克隆性增殖,且從塊莖種植,并生長在粘質土中時,由該植物產生的以鮮重的克數表示的塊莖產量達到每株植物至少大約1000克鮮重的水平。因此,品系agvd17尤其證實使本發明的個體良種品系(即自交可親和的、能育的且基本純合的)互交以得到為本發明的目的的雜種種子的可能性。因此,根據本發明的另一高度優選的植物是品系agvd17的植物或其具有品系agvd17的植物所有生理和形態特征的后代植物。雜種通過如在圖1中例示的本發明的方法產生本發明的雜種植物。通過兩個基本純合的優秀馬鈴薯親本的育種品系的雜交產生本發明的雜種植物,所述優秀馬鈴薯親本的育種品系中包括選自由以下性狀所組成的組中的一些農藝學期望的性狀:昆蟲耐性、線蟲抗性、疾病抗性(包括,但并不限于對粉痂病、絲核菌、銀屑病、晚疫病、由鏈霉菌屬引起的瘡斑病的抗性)、除草劑耐性、耐寒性、耐旱性、耐濕性、耐干腐性和耐濕腐性、耐鹽性、對低溫糖化的抗性和對塊莖脫水的抗性。除了這些農藝學期望的性狀之外,親本育種品系可顯示出如在特定使用市場中期望的那些特征,如期望的塊莖皮的顏色、期望的塊莖形狀、期望的塊莖果肉的顏色、期望的塊莖的味道、期望的塊莖淀粉含量、期望的塊莖干物質含量、期望的塊莖蒸煮品質或期望的塊莖油炸品種。除了這些市場特定的特征之外,親本育種品系當雜交時優選得到顯示出如下有益性狀的雜種:有益的生長率、有益的塊莖產量、有益的塊莖大小、有益的塊莖儲存能力、有益的塊莖休眠期、有益的塊莖保存期限、有益的塊莖制片的品質及有益的塊莖一致性。現在本發明提供了一種馬鈴薯植物的組或一種馬鈴薯植物品系的組,該植物或植物品系是:二倍體(2n);基本純合的(優選在所有同位基因上的純合性超過60%);自交親和的;能育的;及有活力的。迄今已經證明了不可能在單株馬鈴薯植物(品系)中提供所有這些特征的組合。本發明的該馬鈴薯植物的組可以用以產生新型品種或栽培品種。本發明尤其涉及至少兩個二倍體的(2n)、基本純合的(優選在所有同位基因上的純合性超過60%)、自交親和的、能育的且有活力的植物的組,該至少兩個植物能夠雜交以產生根據本發明的雜種品種。馬鈴薯種子育種的新方法新型馬鈴薯植物的供應現在第一次提供了本發明的方法的實現。本發明人已經得到基本純合的、二倍體的、有活力的、能育的且自交親和的,且在它們的基因組內固定有編碼來源于適合的二倍體供體品系的賦予植物農藝學期望性狀的基因的核酸序列的馬鈴薯植物。所述核酸序列是從具有農藝學期望性狀的馬鈴薯種質的基因滲入的結果。傳統上,至少在除了馬鈴薯之外的大部分物種中,原則上可以在僅僅兩個世代中通過已知為雙單倍體(dh)技術的方法使從未受精的生殖細胞(如卵或花粉細胞)得到的單倍體加倍來獲得純合的植物。這在馬鈴薯中是不可能的,因為由于(被認為是)隱性有害基因的表達使迄今所得到的植物示出是不能生殖的。dh方法經本發明人進行廣泛測試,但均發現不能成功。本發明人使用單個自交親和的馬鈴薯品系的馬鈴薯種質和一個或多個含有農藝學性狀的自交不親和的馬鈴薯品系的重復自交、回交和雜交的策略。通過二倍體馬鈴薯植物的持續自交且選擇能育的后代(除了對自交親和的固定選擇之外)來獲得本發明的純合植物。這完全是一種新的途徑,因為在現有技術的方法中,是通過選擇農藝學性狀(如后代植物塊莖形成或一般生長特征)而不是通過從自交中選擇能育的后代來獲得植物。此外,與現有技術的方法不同的是,該程序是使用二倍體植物品系而不是使用四倍體植物進行的。該完全新的途徑意外地證實能夠成功地獲得純合性增加的二倍體后代植物,且該獲得的后代植物是自交親和的且是能育的。而且,當與含有有益性狀(如抗性或白色塊莖顏色)的二倍體品系的植物(異型)雜交時,這些性狀可以被引入基本純合的種質中,且不會丟失自交親和性和能育性。用于獲得所述植物的方法基本如下:起始點是二倍體自交親和的馬鈴薯品系(適合的品系的實例已經被保藏在保藏編號agvd1、agdv2、agdv3和agvd17下)。由于存在sli基因而自交親和的品系通常可被用作起始點。該二倍體自交親和品系的植物自交,且從攜帶種子的漿果中獲得的種子被播種且長成后代植物。攜帶花的后代植物再次自交,且從攜帶種子的漿果中獲得的種子再次被播種且長成后代植物。沒有(或至少很少)注意到在這一階段的植物的農藝學外貌。它們可能開上去是干枯和脆弱的,僅有少量的花。但是,自交至少重復三次,但優選是四次或五次或甚至六至八次。通過這種方式,基本通過自交和選擇產生能育后代的植物來獲得基本純合的、能育的、自交親和的、二倍體的植物品系。接下來,通過使該品系的植物與無關聯的基本雜合的二倍體馬鈴薯品系、優選包含農藝學期望性狀的品系的植物雜交,將農藝學期望的性狀加入基本純合的、能育的、自交親和的且二倍體的馬鈴薯基因型中。該基本雜合的品系(該術語僅僅反應不是如本文使用的基本純合的品系的品系)起到農藝學期望性狀(在這種情況中是活力)的供體的作用。得到的雜種自交和/或回交至自交親和的品系,同時選擇較高的活力。這樣,增強了基本純合的、能育的、自交親和的、二倍體的馬鈴薯植物品系的活力,且同時農藝學上改善了該品系。因此這些步驟將得到有活力的能育的純合自交親和的二倍體馬鈴薯。該活力可進一步通過與其它基本雜合的二倍體馬鈴薯品系雜交來得到增強,該額外的品系用作額外農藝學期望性狀的供體,然后通過自交和/或回交至自交親和的品系,同時選擇這些額外的性狀,該額外的步驟將得到二倍體馬鈴薯育種品系的供應。為了產生優秀的二倍體馬鈴薯育種品系,通過互交、自交和回交,同時持續選擇提供給育種品系的農藝學期望性狀,且通過建立顯示出與其它品系的良好組合能力的品系,來進一步增強育種品系的育種值。通過使用實驗雜交來評估這樣的組合能力。最后,可以通過使顯示出良好組合能力的個體優秀二倍體馬鈴薯育種品系雜交產生雜種馬鈴薯栽培品種。從該純合親本的雜交得到的種子是一致的雜種真正的馬鈴薯種子。使用本發明的方法,通過大約7至8個自交循環將得到基本純合的植物,但技術人員應理解還可使用更多世代的自交。另外,較少的世代(如六個世代)可用以提供本發明的基本純合的植物。回交世代的數量可甚至降低至3,以當應用分子基因標記物時達到超過95%的純合性。產生雜種馬鈴薯栽培品種的新方法基本提供了在大約五年內產生新型馬鈴薯品種的可能性。在實例5中提供了用于從agvd17(f3品系)起始產生雜種馬鈴薯的詳細過程。該品系>80%的純合性,這樣需要非常少量的自交以獲得大約95%~100%純合性的植物。此外,agvd17已經如本文所定義的是有活力的。本發明現在提供了一種用于產生基因同質或一致的真正馬鈴薯種子(tps)的方法。該方法尤其用于在防止病毒感染在馬鈴薯各世代之間傳播的方法中,還用于克服很多與經典馬鈴薯育種、“種子”馬鈴薯生產和運輸和種植相關聯的其它問題。此外,可以以種子的形式保持用于雜種種子生產的當前親本材料,而不是以如通過傳統tps生產所需的植物材料的形式保持。本發明一方面提供了一種用于產生雜種馬鈴薯栽培品種的方法,該方法包括本文中作為起始材料提供的保藏的植物材料的使用。可選擇地,可以開發與本文中提到的保藏不同的其它基本純合的、二倍體、能育的、有活力且自交親和的植物品系,且雜種馬鈴薯栽培品種的生產可以從通過使用本文提供的教導的早期階段水平開始,因為本發明人已經證實該途徑雖然費力,但是成功的。這樣,可選擇的育種材料也是有活力、基本純合、二倍體、能育且自交親和的。作為第一步驟,這樣的早期階段開發可包括提供第一種植的馬鈴薯物種(solanumtuberosum)或相關可交配的物種(通常是結出solanum物種的塊莖)的植物。接下來,所述植物發育成自交親和的二倍體馬鈴薯植物的品系,優選為solanumtuberosum。二倍體馬鈴薯常常自交不親和。可適當地從相關物種引入自交親和性(例如見hermsen,j.g.th.1978,euphytica27,1-11)。然后,自交親和的二倍體馬鈴薯植物的所得品系發育成純合的、能育且自交親和的二倍體馬鈴薯品系。可以通過花藥培養(jacobsen和sopory,1977,theor.appl.genet.52,119-123),通過胚珠培養或通過重復自交(phumichai&hosaka,2006,euphytica149,251-258)通過使單倍體加倍(uijtewaal等.,1987,theor.appl.genet.73,751-78)獲得純合的二倍體。使用后一種方法用于產生形成本文所述的有活力植物的基礎的純合能育自交親和的二倍體馬鈴薯品系。接下來,純合能育自交親和的二倍體馬鈴薯品系發育成有活力的能育純合自交親和的二倍體馬鈴薯品系。可以通過與其它二倍體馬鈴薯植物雜交,自交和回交同時選擇更高的活力來增強活力。必須注意需要高種群,且甚至對于一些用作農藝學期望性狀的供體的無關聯的基本雜合的二倍體馬鈴薯品系,可以在1000株植物的種群中發現非常少量的自交親和的植物。通常,這樣的嘗試結束,且僅當大約10~50%的后代植物是自交親和(如在下面實例2中的d2的情況中)時,供體品系顯示出這樣良好的一般配合力,從而能夠預見到農藝學性狀在自交親和世系中的成功引入。接下來,有活力的、純合的、能育的、自交親和的二倍體馬鈴薯品系發育成二倍體馬鈴薯育種品系。最優選地,產生至少兩種分離的有活力能育純合的自交親和的二倍體馬鈴薯品系,且發育為至少兩種不同的二倍體馬鈴薯育種品系。這樣的育種品系包括大量的農藝學期望性狀。通過與其它二倍體馬鈴薯植物雜交、自交和回交,同時選擇這些性狀來引入農藝學期望性狀。該育種程序可包括測試疾病,如芽或塊莖是否沒有疾病,或它們是否對疾病具有抗性。接下來,二倍體馬鈴薯育種品系發育成優秀的二倍體馬鈴薯育種品系。這使用通過互交、自交和回交,同時持續選擇農藝學期望性狀以增強育種值來完成的。在該階段,進行實驗雜交以評估一般配合力。現在,優秀的二倍體品系還可通過自身已知的方法發育成優秀的四倍體(4x)品系,如通過使用例如秋水仙堿進行化學處理,或通過使用組織培養使基因組“自發”加倍。最后,通過良好的結合育種品系(combiningbreedingline)的雜交,從這些優秀的二倍體或四倍體馬鈴薯育種品系產生馬鈴薯雜種栽培品種。獲得的種子是一致的雜種真正馬鈴薯種子。本發明的用于產生馬鈴薯雜種栽培品種的方法包括產生二倍體馬鈴薯育種品系的步驟。所述方法包括以下步驟:(a)提供第一馬鈴薯植物,所述第一馬鈴薯植物是二倍體、自交親和的且基本純合的馬鈴薯品系的植物,優選是有活力的、能育、純合的且自交親和的二倍體馬鈴薯品系的植物,更優選是二倍體馬鈴薯育種品系;(b)提供可具有任何水平的多倍性(優選二倍體)和/或任何水平的純合性的馬鈴薯品系的第二馬鈴薯植物,其中所述第二馬鈴薯植物用作農藝學期望性狀的供體,所述性狀被基因滲入進入所述第一馬鈴薯植物的所述基本純合的品系中,或至少進入由所述第一植物和所述第二植物之間雜交所得到的后代植物中,其中所述性狀選自由昆蟲耐性、線蟲抗性、疾病抗性(包括,但并不限于對粉痂病、絲核菌、銀屑病、晚疫病、由鏈霉菌屬引起的瘡斑病的抗性)、除草劑耐性、發根力、耐寒性、耐旱性、耐濕性、耐干腐性和耐濕腐性、耐鹽性、生長率、塊莖發育缺陷(例如畸形或受損塊莖)、塊莖產量、塊莖大小、塊莖皮的顏色、芽眼深度、塊莖形狀、塊莖肉的顏色、塊莖味道、塊莖儲藏能力、塊莖休眠期、塊莖保存期限、對塊莖脫水的抗性、塊莖淀粉含量、塊莖干物質含量、塊莖蒸煮品質、塊莖油炸品質、塊莖制片的品質、塊莖的一致性和對低溫糖化的抗性所組成的組中。在提供了這兩種植物之后,該方法又包括以下步驟:(c)所述第一馬鈴薯植物和第二馬鈴薯植物異花授粉,以便所述親本植物中的一種親本植物產生攜帶種子的果實(漿果),且從所述果實中收集所述種子以提供雜種后代的種子。然后,種子長成植物,且直接或在額外的自交和回交步驟之后從供體植物中選擇包括農藝學期望性狀的植物,如在植物育種領域中公知的。在這些方法中,可以使用標記物以輔助育種過程。雜種后代種子優選長成植物,該植物隨后與植物回交至回歸親本品系。后面的步驟對于上述方法中的任一種不是必需的。但是,這些額外的步驟可以通過使本發明的種子生長提供消費塊莖的產生,和/或可提供新型純合、自交親和的二倍體優秀育種品系的產生,該新型純合、自交親和的二倍體優秀育種品系可用作使進一步的性狀包含進入后代中的親本。本發明的植物是能育的和有活力的。這些額外的益處通過下面進一步的步驟獲得:(d)使所述后代種子長成雜種后代馬鈴薯植物,且(e1)從所述雜種后代馬鈴薯植物中收獲塊莖以提供攜帶所述性狀的塊莖,或(e2)使所述雜種后代馬鈴薯植物自交或使所述雜種后代馬鈴薯植物與所述第一馬鈴薯植物的所述品系的馬鈴薯植物回交,以提供進一步的攜帶所述性狀的后代植物,且其中所述后代植物在每次回交或自交步驟之后純合性增加。優選地,在如上所述的方法中使用的所述第二馬鈴薯植物是如在上面步驟(d)中定義的雜種后代馬鈴薯植物,且通過如上所述的方法獲得。更優選地,在如上所述的方法中使用的所述第二馬鈴薯植物是如在上面步驟(e2)中定義的雜種后代馬鈴薯植物,且通過如上所述的方法獲得。優選地,所述步驟(e2)包括2~7次回交,使用品系agvd1、agvd2、agvd3或agvd17的植物作為回歸親本,且測試后代植物中性狀的存在(例如造成性狀的等位基因以純合或雜合形式存在),然后通過一次或兩次(或更多次)自交,以提供基本純合的植物,該基本純合的植物在基因方面相當于種群agvd1、agvd2、agvd3或agvd17的植物,但其中造成性狀的等位基因雜合存在,優選純合存在。該植物具有至少一個在種群agvd1、agvd2、agvd3或agvd17中沒有的額外的等位基因。現在上述方法提供了一種用于基于種群在種群agvd1、agvd2、agvd3或agvd17的背景產生純合育種品系或優秀育種品系的育種過程,在該背景中先后加入額外的等位基因,從而提供其中堆疊有期望性狀的優秀的育種品系。本發明的方法現在第一次提供了一種用于防止病毒感染在馬鈴薯世代之間傳播的方法,同時還解決了現有技術中很多其它與新型馬鈴薯品種育種和消費用馬鈴薯的商品化生產相關的問題。通過提供真正馬鈴薯種子(即植物學種子)使得該方法成為可能,該種子可以長成產生用于消費的塊莖的馬鈴薯植物,且該種子來自至少一個親本是二倍體、自交親和且基本純合的馬鈴薯品系的雜交中,優選所述種子來自其中兩個親本均是二倍體、自交親和且基本純合的育種品系的雜交中。該新方法的重要優點是以這種方法病毒感染的傳播不會通過配子體散布,且種子可以有效地防止被病毒感染。本發明人現在提供了一種用于進一步培育容易操作且顯示出改善的能育性和植物活力的純合品系的方法,當與現有技術的近交相比,該方法同時維持自交親和性和高水平的純合性。應理解正如在番茄中一樣,并總是需要100%的純合性以提供有效的育種程序。只要維持自交親和性,則這不是問題。現在,已經第一次成功示出農藝學期望或有益的性狀進入純合馬鈴薯品系中的基因轉移。基于本文提供的手段和方法,技術人員應能夠基于他自己的種質成功產生純合的馬鈴薯育種品系。此外,技術人員應理解他通過提供的手段和方法能夠例如將額外的性狀堆疊進入目前提供的自交親和和基本純合的品系中,以建立富含農藝學期望性狀且可在他們的權利中的商品化育種中用作種質的品系的種群。實例實例1.育種程序和譜系的一般描述作出很多嘗試以產生馬鈴薯的純合、能育、自交純合的二倍體基因型,但沒有成功。這些嘗試包括使單倍體加倍,及隨后篩選能育花的形式。利用了幾種方法,像花藥培養以再生單倍體且最終使單倍體純合植物加倍、重復自交以逐漸增加在現存的二倍體種質中的純合性水平,且通過使用s.phureja作為花粉供體進行皮刺授粉(pricklepollination)以誘導從未受精的卵細胞形成種胚。特別是通過分析超過兩百萬的后代大規模地實施了后一種方法,但沒有獲得期望的成功。這些測試沒有得到有活力的自交親和的植物。通過來源于公開的種質的品系的植物多個世代的自交獲得基本純合的馬鈴薯品系,該植物具有控制自交親和性的sli基因(phumichai等.euphytica(2006)148:227–234)。這些植物(在本文中被定名為“h”或ivp07-1001/4)生長不良,具有非常少量的花且每個漿果僅包含少量的種子。此外,上述植物非常小(最大60cm)。這些植物幾乎不產生塊莖,且形成的塊莖也非常小(平均塊莖重量大約為5克/塊莖)。總之,這些植物沒有用作商品化馬鈴薯育種的價值,如它們顯示出植物重量和塊莖產量小于二倍體育種克隆rh89-039-16的20%。上述植物僅在它們的基因組中的很多基因座上含有純合的等位基因,是自交親和的且是二倍體。在2008年的夏天,該ivp07-1001/4品系的植物與不同的二倍體供體品系(d1、d2和d3,見完整譜系圖2)雜交。d1是早期,具有長尺寸的橢圓形塊莖、淺黃色果肉和良好蒸煮油炸制作品質和良好的蒸煮品質,且用作雄性親本(表6)。得到的子代在該實例中被稱作f1(d1)。以相似方式但使用供體d2作為雌性親本獲得其它f1后代植物。d2是基本非純合但不是自交親和的優秀的二倍體育種克隆,但d2具有早期蒸煮、橢圓形塊莖、黃色果肉和制片的良好品質。這樣得到被定名為f1(d2)的品系。上面的是d1的f1自交以產生f2(d1)。該品系被保藏在培植者編號agvd1下。估計agvd1為大約50%的同源性,意味著致使親本不同的基因座中有大約50%是純合的。f1(d1)與f1(d2)雜交以產生偽f2。該品系(f2(d1/2))被保藏在培植者編號agvd2下。估計agvd2含有大約25%的源自ivp07-1001/4的同源等位基因。是d1的f1還與ivp07-1001/4回交以產生bc1品系。該品系(bc1(d1)和bc1(d2))被保藏在培植者編號agvd3下。估計agvd3對于致使親本不同的基因座為大約75%的同源性。通過使用供體d3作為雌性親本以相似方式獲得其它f1后代植物。d3是具有黃色果肉且具有抗性基因r3、h1、gpa2、rxadg的二倍體育種克隆。這樣得到定名為f1(d3)的品系。與上面描述的相似,f1(d3)自交以產生f2(d3),且與sli基因供體回交以產生bc1(d3)后代。f1(d1)與f1(d2)雜交以產生偽f2。該品系(f2(d1/2))被保藏在培植者編號agvd2下。估計agvd2對于致使親本不同的基因座為大約25%的同源性。f1(d1)還與ivp07-1001/4回交以產生bc1品系。該品系(bc1(d1))被保藏在培植者編號agvd3下。估計agvd3對于致使親本不同的基因座為大約75%的同源性。bc1在冬季苗圃中自交,得到bc1s1種群。下一個夏天(2010年),f2、偽f2和bc1s1被播種在苗圃中的苗床上,且被移植在小的泥炭盆中。在2010年6月,幼苗被移植至荷蘭的粘土和沙質土壤處的大田中。總計從幼苗生長出超過15000株植物,且對這些植物選擇自交親和性和良好的塊莖品質和產量。大部分f2和bc1s1種群示出非常低頻率的自交親和性。選擇具有合理的良好漿果和結實率的罕見植物,用于子代測試,而不進行進一步地農藝學評價。僅有兩個源自d2xh雜交的f2種群示出大約25%的自交親和性。對于自交親和最強的種群的每一個種群,選出最多15株具有最佳塊莖品質和數量的植物用于子代測試。偽f2的植物比其它種群示出更強的活力,可能這些植物不含有任何具有供體親本的純合等位基因的基因座。這些種群中的一個種群示出超過50%自交親和性。與上面描述類似,還選出自交親和最強的種群中的最多15株植物用于子代測試。此外,在供體d1~d19的兩株或十二株植物(實例3)上,進行與來自大田作為花粉供體的自交親和的植物雜交。在評價塊莖品質和數量之后,如果缺少足夠數量的雜交,則僅選擇與所選擇的植物或它們的全同胞(fullsib)雜交的用于子代測試。這樣,在每一世代中,選擇自交親和且具有良好農藝學性能的植物。為了產生純合的品系,收集這些所選擇植物的自交漿果的種子用于進一步地測試。此外,所選擇的植物與其它二倍體供體或其它育種品系雜交以增強基因變異。這些f1種子是雜合的,且需要幾輪近交以產生具有期望性狀的純合的后代。這樣,每年可以進行兩個世代的選擇、自交和雜交。甚至通過使用對特定性狀的診斷標記物及隨機標記物以選擇回歸親本的基因組,利用標記物輔助育種增強該過程。當已經獲得優秀親本時,可以通過兩代至三代的回交及兩代自交制成這些品系的衍生物。實例2.培植自交親和的二倍體馬鈴薯1.引言二倍體馬鈴薯的共有特征是自交不親和性(si);當自受精時不會產生有生機的子代。已經描述了源自solanumtuberosum的s-基因座的抑制基因(sli)(euphytica99,191-197,1998)以抑制該自交不親和性,且使二倍體馬鈴薯變成自交親和(sc)。當重復自交時,已經產生接近純合的自交親和的二倍體馬鈴薯克隆(genome48,977-984,2005)。這些克隆的農業學品質非常差,實際上反映了不良的萌芽、植物生長、塊莖品質和塊莖產量。2.實驗2.1材料為了開發具有更高水平的農業學性狀的自交親和的馬鈴薯克隆,我們使sli基因供體與定名為ivp97-079-9、ivpaa-096-18和sh83-92-488的不同的馬鈴薯克隆雜交。這些克隆可以在r.c.b.hutten博士(植物育種實驗室,瓦格寧根大學,荷蘭)處獲得。如在本文在所使用的,這些克隆被分別定名為克隆d1、d2和d3。子代(f1)自交且與sli基因供體回交,分別產生f2和bc1種群。bc1自交以產生bc1s1種群。在除了植物的自交親和性之外沒有任何其它的選擇的情況下,進行所有這些雜交和自受精。2.2在冬季試驗中對子代的評價在預實驗(pilotexperiment)中,幾個隨機的bc1和f2子代生長在熱的溫室中達接近三個月,該熱的溫室具有16小時人造光,而外部溫度低于零度。我們以人工授粉之后具有漿果的植物的數量評價了自交親和的百分比,并以每株植物的塊莖的重量評價了產量(表2)。結果sli供體是自交親和的(sc),且大部分f1和bc1植物也是自交親和的,而少數f2植物是自交親和的。sli供體示出極低的塊莖產量,而bc1和一個f2是f1塊莖產量的大約50%。一些bc1植物的塊莖產量接近f1植物的塊莖產量,且一些(未選擇的)f2植物好于最弱的f1植物。表2.二倍體馬鈴薯育種程序以引入sli基因的冬季生長植物的一些特征*來自塊莖**來自幼苗2.3大田中子代的評價在2010年的夏天,在大田中測試f2和bc1s1種群的子代植物以選擇具有良好農業學性能的自交親和的子代植物(表3)。表3.將自交親和性(sc)引入二倍體馬鈴薯中的育種模式。粗體指示為最有希望的材料。*足夠的結實率結果我們測試了包括30至840株植物的52個f2種群,及包括1至390株植物的62個bc1s1種群,總計分別有9660株f2植物和3901株bc1s1植物。大部分種群示出弱的植物生長和低頻率的自交親和性。僅有來源于d2親本的兩個f2種群示出大約25%的自交親和的植物。基于塊莖產量和塊莖品質,選擇這些種群中的最佳植物。3.結論在52個f2種群中僅有兩個種群且沒有bc1s1種群示出合理水平的自交親和性。但是,可以從要不然已經被培植者忽略的材料起始出人意料地獲得期望的植物。本發明的植物的與現有技術的那些植物的主要不同給出在下面的表4中。表4.建議用于產生雜種(真正)馬鈴薯種子的本發明的親本馬鈴薯品系的特征。實例3.分子標記物平臺的開發引言能夠獲得很多標記物系統以支持基因研究。對于本發明,開發了馬鈴薯中的分子標記物平臺,且將該分子標記物平臺用于評價二倍體馬鈴薯中的純合性比雜合性的水平。用于snp檢驗的大部分提供信息的序列的選擇為了分析馬鈴薯中的snp,之前已經使用了illuminastandardgoldengate方案(shen等.2005.mutationresearch573:70–82)(anithakumari等.2010.mol.breeding,26:65-75)。該平臺包括384個snp,且后面使用768個snp進行擴充。已經使用est序列開發了這些384個和768個snp,est代表沒有內含子序列的mrna。由于本發明人已經獲取了馬鈴薯基因組的第一序列草圖(www.potatogenome.net),因此能夠將mrna與包含間插序列的基因組dna序列進行對比。使用本地序列基本搜索工具(basiclocalalignmentsearchtool)(blast;altschul等.1990.j.mol.biol.215:403-410)進行該與馬鈴薯基因組序列的對比。這樣分別得到538和1681個搜索結果(hit)。多個blast搜索結果的出現被認為反映間接同源(paralog)或內含子跨越序列(intronspanningsequence)。將這些搜索結果從為goldengate分析首次開發的snp初始集合(anithakumari等.2010)中去除。得到279個和453個唯一的blast搜索結果,代表迄今已知的solanumtuberosum的phurejadm1-3516r44(cip801092)組的馬鈴薯基因組序列(馬鈴薯基因組測序協會(potatogenomesequencingconsortium),www.potatogemome.net)中僅有的一個拷貝的出現。從具有唯一blast搜索結果的總共732個snp中,由于高水平的序列錯配(101bp內有3~6個snp)僅排除少量的snp(31個事例,平等地分布在384和768的組(set)中)。應預期一個錯配,且容忍第二個snp以將其包含在所選擇的snp-跨越序列中。對于現在的snp設計,將最小的序列長度設置為101bp。因此,81個序列因為它們較短而被去除。得到的553個序列具有至少101bp的長度。然后因為在經加倍的單倍體的支架(scaffold)內具有僅35個直到94個核苷酸的不完全匹配而去除總共100個序列(dm-3516r44;www.potatogenome.net)。還去除了具有引入的對比空位(alignmentgap)的序列及具有其它潛在錯配問題的序列。雖然有25個序列沒有滿足上述標準,但仍然保留了它們,因為這些序列已知在goldengatesnp檢驗中是有高度信息量的。這些snp甚至允許區分在四倍體(voorrips等,eapr-eucarpiacongress“potatobreedingaftercompletionofthednasequenceofthepotatogenome”,27-30june2010,wageningen,thenetherlands,e-publicationathttp://edepot.wur.nl/143559,pg42)中出現的五種基因類別。最后,選擇出515個序列。最終根據馬鈴薯基因圖譜上的(515個標記物中的)237個標記物的已知圖譜位置選擇出100個序列(anithakumari等,2010和vanos等,genetics173:1075-1087,2006)。這些選擇的標記物均勻分布在馬鈴薯12條染色體上,包括最遠端端粒以及著絲粒標記物,該選擇的標記物對在克隆rh(rh89-039-16;www.potatogenome.net)中是雜合的snp有偏向性,因為克隆rh是本研究中使用的材料的祖先。或者換句話說,克隆rh的等位基因也存在于在本研究使用的材料中(見表2和圖3)。標記物的測試如上所述的選擇的100個snp序列用于kasparsnp基因分型系統(kbioscience有限公司,hoddesdon,uk:http://www.kbioscience.co.uk/lab%20services/snp%20genotyping/genotyping_chemistry.html)。在包括用于本研究的三種二倍體親本的20種二倍體馬鈴薯基因型的組(見圖4)上測試標記物,且進一步在sli基因的供體(克隆07-1004-1)(同樣參見本文中的實例2)上測試標記物。結果所有snp序列均在kasparsnp基因分型系統(見圖4)中產生多態性,對應于100%的成功率。總共測試了100種標記物x20種基因型,包括2000個數據點。我們觀察到有29個缺失數據(低于2%)。作為對照,對一種基因型測試兩次,據此使用不同位置的植物樣本(圖4,第4列和第7列)。所有數據都相同。這些結果表明kasparsnp基因分型系統是一種用于二倍體馬鈴薯的有效、可靠且可重復的標記物平臺。這也表明kasparsnp基因分型系統對二倍體馬鈴薯來說是有效、可靠且可重復的。所有標記物均示出在該實驗中使用的二倍體馬鈴薯種質的出現頻率較高(附錄iii)。通過要求這100種標記物使得種質中的雜合子頻率在26%至77%的范圍內變化,同時sli基因供體為100%的純合性,這與期望值一致(phumichai等,2005genome48:977-984)。在本研究中使用的sli供體以及三個親本之間的提供信息的標記物的數量分別為54、52和62。這表明kasparsnp基因分型系統同樣能夠為二倍體馬鈴薯提供信息。基于上述情況,得出的結論是本文所述的基于本文開發的標記物的kasparsnp基因分型系統對描述二倍體馬鈴薯中的純合性/雜合性來說是有效、可靠、可重復的,且能夠提供大量信息。表5a.馬鈴薯100種snp標記物的序列表5b.potsnp標記物及其在圖3的sh和rh圖譜上的位置注釋的列表表6用于snp標記物開發和測試的二倍體馬鈴薯基因型早熟:早期開花(周期短);qcook:良好的蒸煮品質;qfry:良好的油炸品質;phyt表示晚疫病抗性基因實例4.純合二倍體馬鈴薯植物的開發及雜合f1雜種子代的生成引言該實例的目的在于開發自交親和性(sc)且具有良好農藝學性能的純合二倍體馬鈴薯品系。上述實例2中描述了以結合自交親和性以及良好農藝學性能的育種程序。本次實驗的目的在于證明可通過近交生成純合的二倍體自交親和的馬鈴薯基因型,且包含散布在馬鈴薯基因組上的相對的純合的基因座的二倍體馬鈴薯植物可以進行雜交(互交)以生成雜合f1雜種子代。材料與方法在上面實例中描述的含有sli-基因的自交親和的二倍體純合的馬鈴薯基因型與定名為d1和d2的兩個馬鈴薯克隆雜交(見實例1和2)。f1子代自交,且由兩株隨機的scf1植物生成兩種f2子代。這兩個f2種群的隨機自交親和植物自交,且對于每個f2種群,測試三個f3種群的純合性水平以及snp標記物的分離。測試6株f2植物的倍性水平(通過dna的熒光定量),且發現該6株f2植物都是二倍體。應用涵蓋馬鈴薯所有24條染色體臂(見圖5)的kasparsnp基因分型系統(見實例3),將一組36個標記物用于265株個體f3植物上。結果在總共265株植物中,對于每對親本對使用至少24種提供信息的標記物測試來自六個f3種群的35~56株植物的子集。總共使用了36種標記物(見圖5)。標記物在馬鈴薯基因組上的位置見上述實例3。sli供體對測試的這些標記物來說是100%純合的,同時對于每對親本對,d1和d2親本在24種提供信息的標記物中分別包含雜合的20和19個基因座,而其他五個和六個基因座包含來自供體親本(d1或d2)的純合等位基因。六株純合的f3植物如所預料的那樣,f3種群因f2中雜合的標記物而不是因在f2中固定的標記物而分離(圖5)。平均雜合子水平在84%至95%的范圍,同時個體植物的雜合性在71%至100%的范圍內(表7)。表7.六個f3種群的純合子頻率(%)f3種群供體親本平均值范圍126d18574-96126bd18874-100127d18471-96121d28571-96122d28979-100123d29488-100使用超過100種提供信息的標記物確定從育種程序選擇的一組f2、f3和f4植物的表型和基因型。根據自交親和性、良好植物性能(活力)和良好塊莖品質和產量來選擇這些植物。通過應用如上所述的kasparsnp基因分型系統進行基因分型(見:用于snp檢驗的大部分提供信息的序列的選擇;表5)。>100種標記物均勻分布在12條馬鈴薯染色體上。這樣提供了比上面使用24種標記物甚至更好的覆蓋率。在重復試驗中,f2植物的產量在50克/植物至2500克/植物的范圍,同時一些其它二倍體雜交得到在對照四倍體栽培品種的平均值之上的高達3100克/植物的產量。如在表9中所示,基于圖5中的數據,分析d1xh及hxd2雜交得到的三個f3種群的純合基因座的頻率。期望這些頻率給出親本等位基因的頻率。三個種群中有六株植物已是100%純合的。這些植物主要含有具有sli供體等位基因的純合基因座。這被認為是sli供體含有純合等位基因的緣故,同時對于每個雜合的標記物而言,其他親本包含一個與sli供體共有的等位基因。所以,來源于sli-供體的等位基因在全部后代種群中都很豐富。被定名為122-34、123-6、123-20、123-23、123-24和126b-17的這六株純合f3植物分別包含具有來自供體親本的純合等位基因的兩個、五個、四個、三個、五個和七個基因座。所以,代表24條染色體臂的24種標記物基因座已經在兩個自交世代中達到100%的純合性。類似地,在選擇的f2、f3和f4植物上使用>100種標記物的基因分型數據示出個體植物的純合性百分比的增加,f2的27~70%至f3的58~87%至最高純合性的f4種群的85~99%。無絕對致死的等位基因常有報告指出二倍體馬鈴薯中的重復自交由于近交衰退緣故只會產生衰弱的植物。這應歸因于純合存在的降低植物適合度的大量有害等位基因。因此,我們在提到的六個f3種群中測試了36種標記物的分離(見圖5)。還測試了這些標記物在19種選擇作為育種程序的供體的二倍體馬鈴薯基因型中的頻率(見實例3,圖4和表9),選擇這些標記物是因為它們能為本研究中的親本對提供信息。如果存在致死基因,則不應存在具有純合等位基因的基因座。實際上,七種標記物(定名為potsnp165、potsnp205、potsnp753、potsnp238、potsnp700、potsnp100及potsnp607)沒有示出不同于該二倍體種質中的sli-供體等位基因的純合等位基因。這些標記物大部分已固定在f2中,它們中的大部分因sli供體等位基因而在親本中更加豐富,但同樣觀察了固定有針對非sli親本的等位基因(種群126b和127中的potsnp753)的f3種群的實例。這七種標記物中的三種(potsnp205、potsnp753及potsnp100)在f3種群中分離,從而potsnp753正常分離,而potsnp205及potsnp100表現為偏(distorted)分離。這些可代表有害等位基因。只鑒定出一種具有純合potsnp100供體的等位基因的植物。在不希望受理論約束的情況下,認為這可能代表非常有害的等位基因。由于已經鑒定出單株具有供體親本的純合等位基因的植物,因此不能充分證明為絕對致死等位基因。f3×f3雜交2010年的夏天,將測試的265株f3植物種植在不加熱的溫室中。此外,將大約600株另外的f3植物種植在粘土土壤的大田中(同樣見實例2)。所有這些植物均來自荷蘭冬季苗圃中生長的未經選擇的f2植物(見實例2)。通過在溫室生長的自交親和的f3植物的柱頭上涂覆來自溫室及大田生長的自交親和植物的花粉來進行雜交。僅在具有不同曾祖代親本的f3植物之間進行雜交(種群121-123的f3植物與種群126-127的f3植物雜交)。定名為126-24、126-31、126b-2及127-21的溫室植物作為雌性自交親和親本與來自種群123的自交親和f3植物作為雄性親本成功雜交。f3雌株的純合性大于80%,而不測試大田中的f3植物,但種群平均純合性水平為94%,范圍在88%至100%之間。對比f3xf3雜交的親本中已固定在f3種群中的基因座(見表8)。鑒定出在九條染色體上的十個純合基因座,該十個純合基因座在用于f3xf3雜交的四個雌性親本中的一個和一個雄性親本之間至少有明顯差異。因此,f1雜種子代對于這些有相對(contrasting)的基因座來說至少是雜合的。由于每一個體親本對包含至少五個有明顯差異的基因座,因此每個個體子代植物將含有至少五個有明顯差異的染色體區域。表8.對比f3xf3雜交的親本之間的固定的基因座代碼126-24、126-31、126b-2及127-21是指在與定名為“123”(見上文)的f3種群的一個大田生長的植物雜交中用作母本的個體f3植物。按照純合基因座占所有測試基因座的百分比計算純合性水平(圖5)。“x”是指在四株雌性植物及種群123的一株植物之間的明顯不同的等位基因的純合標記物。結論發現自交兩代后,獲得純合的二倍體馬鈴薯植物,包括來自兩個親本的等位基因。測試的36種標記物中沒有一個標記物提供致死等位基因的證據。此外,具有大于84%純合基因座的植物可成功雜交以獲得具有至少五個雜合染色體區域的雜種植物。這意味著現已經生產出了基本上一致的且代表真正馬鈴薯種子的雜種種子。表9.在二倍體種質和f3種群中的全基因組snp標記物的等位基因頻率#:植物的數量;hh:純合的sli-供體基因座;dd:純合的供體基因座;hd:雜合的基因座;%是雜合植物的百分比,數量小表示f3中的純合性水平高。實例5.商品化馬鈴薯品種的育種模式從上述實例2和4中所述的sli供體和另一二倍體馬鈴薯之間的雜交獲得的f3材料起始,開始下面闡述用于生產商品化馬鈴薯品種的育種模式。該模式由多個連續步驟組成,結合本文所述教導,允許技術人員獲得與現有商品化雜種一樣優質、或更優質,且在五年內比傳統育種品種繁殖快得多的二倍體或四倍體的雜種馬鈴薯品種。1.產品品種定義根據本文所述的本發明,基于商品化雜種的生產,新型馬鈴薯品種的生產并不是機遇問題,而是一個由該品種產品的連接起始的直接設計的過程。本文中,必須定義用于特定市場(馬鈴薯片、淀粉產業、消費的馬鈴薯)的期望產品的概況(目標產品概況)。此外,必須定義產品概況中對相關性狀的評價方法。優選地采用診斷標記物評價相關性狀的存在。這樣的標記物在傳統馬鈴薯育種領域是熟知的。2.大田試驗根據開發過程開始的季節,選擇合適的試驗田。當在九月或十月開始時,優選選擇位于南半球的合適的試驗田。大田試驗可起始于至少2個、優選更多的f3種群,所述f3種群選自二倍體sli供體與另一充當農藝學性狀供體、優選活力性狀供體的二倍體馬鈴薯基因型雜交種的至少一個、但優選多個,其中f3種群代表如本發明的實例2中例示的二倍體、能育的、自交親和且基本純合且有活力的馬鈴薯品系。除了這些f3種群之外,f1子代取自至少兩株,優選多株經選擇的f2植物(如上述實例2中所例示的),所述f2植物與經選擇的f2植物的祖代不同的至少兩種、優選多種二倍體馬鈴薯基因型雜交。然后讓這些f3和f1種群的至少50株、優選更多的個體植物在選擇的馬鈴薯大田中生長,且自交。3.評價與選擇在大田中生長的過程中,對開花后2~6周之內帶有漿果的自交親和植物的頻率進行評價,且選擇具有至少10%、優選更多的自交親和植物的至少兩個、優選更多的f3和f1種群。進一步地,在生長季節中,對所選種群中的自交親和植物的產品概況中所述的農藝學特征進行評價。采集所選種群的所有自交親和植物的葉片樣本用于dna標記物的測試。在生長季節結束時,對所選種群中的自交親和植物的塊莖品質和數量進行評價,且利用診斷且隨機的標記物對dna標記物基因型進行評估。基于該評價及評估,在下列條件的基礎上在每個種群中選擇至少五株、優選更多的自交親和的植物:(i)最接近目標產品概況的性狀組合;(ii)最高純合性水平;及(iii)最大數量的相對純合的基因座。經選擇并自交的f3和f1植物分別提供種子形式的f4和f2子代。4.塊莖和種子的收獲及運輸開花后在至少六周、優選六周以上的時間段內收獲所選的f3和f1植物的塊莖以及這些所選植物的漿果中的f2和f4種子。采集f4種子用于進一步的包括測試由此長出來的f4植物之間的一般配合力的大田試驗,且f2種子為育種提供額外的基因庫,并采集f2種子用于進一步的大田試驗。根據季節,且為了保證時間,可適當將種子運輸至北半球以在溫室中生長,但條件是要滿足所有涉及的國家植物檢疫法規。5.塊莖儲藏及評價所選的f1和f3植物的塊莖優選儲藏在種植地區以免不希望的病原體和害蟲的傳播,且評價這些儲藏塊莖的在產品概況中定義的儲藏塊莖的某些相關內在的塊莖性狀。6.1年后塊莖的增殖及評價在下一季種植所選的f1和f3植物的塊莖,優選種植在與原來馬鈴薯田相同的區域,以免不希望的病原體或害蟲的傳播。每個地塊使用至少兩種、優選更多種馬鈴薯塊莖,且每一克隆優選使用一個、優選多個復制本。使用至少一種、優選多種有關的商品化栽培品種作為對照。優選使用具有商品化栽培品種的邊行(side-row)以避免邊緣效應。對產品概況中定義的有關植物特征(例如蒸煮、油炸、制片、淀粉含量等)進行評價。進一步評價每個地塊的塊莖產量以及如在產品概況中定義的內在相關的塊莖特征。這樣的評價用于家族特征化及子代選擇。每年都重復進行這些試驗,優選在南北半球位置都進行以便連續選擇在大田中生長的植物的祖先性質。在生長后期,當獲得雜種時,進行商品化引入的測試,將評價試驗推廣到針對在至少一平方米、優選多個平方米的地塊大小上的至少兩個、優選更多個復制本,且推廣至至少兩個、優選更多個位置上進行,但不限于目標市場。7.與f4種子測交及大田試驗采集所選的自交f3植物的f4子代種子之后不久,播種這些種子中的至少五十、優選更多個種子。如果需要,可以在加熱溫室中進行該播種。播種后大約1個月后優選將幼苗移植到小盆(pot)中,然后在幾個星期之后被移植到合適位置的大田中。繼續進行上述評價和選擇,且每個種群中的至少兩株、優選更多株所選的f4植物與具有最大數量的相對基因座的兩個、優選更多個所選親本的至少兩株、優選更多株其他的所選f4子代植物測交,以生產測試雜種(f4xf4→雜種)。另外,每個種群中的至少兩株、優選更多株所選f4植物與具有所選植物所沒有的期望特征或性狀的至少一種、優選更多種的二倍體基因型雜交,以提高育種種質的基因多樣性。進一步地,每個種群中的至少兩株、優選更多株所選f4植物自交以生產f5種子。更進一步地,對每個種群中的至少兩株、優選更多株所選f4植物的植物的一部分進行滅菌,體外增殖,并用合適的化學品(秋水仙素/俄瑞扎林(oryzaline))進行處理,以產生四倍體f4基因型,且利用四倍體f4植物在與上述相同的親本之間進行測交。每年重復進行上述育種模式,由此實現了產品概況中定義的植物性狀的持續提高。由測交得到的塊莖的評價有助于確定親本的一般配合力。一般配合力在本文被定義為一系列雜交中品系的平均性能。針對每個市場實現至少兩個、優選更多個產生商品化雜種種子的優秀親本品系。預計在五年內生成與現有商品化雜種一樣優質或更優質,且比傳統育種品種增殖快得多的二倍體或四倍體雜種品種。實例6.期望性狀的引入為了產生根據本發明的新型雜種品種,產生基本純合的育種品系,從該育種品系可以通過兩個基本純合的親本育種品系衍生出f1商品化雜種。這些基本純合的親本育種品系是由一致的二倍體、自交親和且有活力的植物組成的品系,該一致的二倍體、自交親和且有活力的植物具有每株植物多于200克、優選500克的新鮮塊莖產量。依賴于親本的一般配合力,這些親本品系的每一株均攜帶被轉移入后代f1中的性狀。親本品系是本發明的一個方面。相對于在早期共同待決的申請pct/nl2010/050716中描述的植物,這些植物包括額外的性狀。這些額外的性狀可包括任何農藝學期望的性狀,優選地選自在表10中提供的性狀。為了產生本發明的親本品系,可使用下面的方法。a.從已知來源并入農藝學期望的性狀在性狀已知,但唯一的可用信息是關于可用作性狀來源的供體加入物(accession)或供體品種的知識的情況下,可適當地使用下面的方法以產生本發明的植物。提供包括期望性狀的(野生型)加入物或品種的植物,且在所述性狀供體品系的植物和來自基本純合的育種品系的植物(本文指受體育種品系的植物)之間進行雜交。根據本發明的基本純合的育種品系是二倍體、自交親和、有活力、基本純合且具有相當于超過200克、優選超過500克新鮮塊莖重量的塊莖產量的品系,在該品系中引入了額外的期望性狀。從通過上述雜交產生的雜種后代植物中,通過進行對性狀存在進行表型篩選(如生物檢驗)選擇顯示出所述性狀的植物。使用基本純合的受體育種品系作為回歸親本,且再次通過使用表型篩選選擇顯示出所述性狀的后代植物,進行重復回交以監控導致期望性狀產生的基因從性狀供體至受體育種品系的成功轉移,其中所述回交步驟用以保持受體育種品系的基因組背景。進行重復的回交和選擇過程,以便留下供體的不期望的性狀而使感興趣的基因轉移至受體育種品系中。然后,這些步驟提供在其基因組中收集有農藝學期望性狀的額外基因的馬鈴薯育種品系。最后,從這些回交和選擇步驟得到的植物自交以提供具有并入其中的期望性狀的純合的受體育種品系。在性狀遺傳(顯性對隱性;單基因對多基因)上的基因信息是已知的情況下,可以對回交和選擇步驟進行優化以使性狀從供體轉移至基本純合的受體育種品系中。技術人員應意識到需要各種步驟以優化該轉移。例如,在隱性性狀的情況中,為了確定雜種是否包含以雜合形式存在的性狀的等位基因,表型篩選應涉及雜種的額外自交。包含等位基因的植物用于進一步地回交。在包括期望性狀的來源加入物或品種不能與來自基本純合的受體育種品系的植物雜交的情況下(即如果它們是不可雜交的物種),可使用下面的用于種胚挽救的方法。一株植物的花粉用以使另一植物的胚珠受精(其中任意一株可以是供體或受體),受精后形成種胚,且通過從胚珠分離來挽救得到的種胚。可以使合子種胚與它們的種皮分離,然后被能夠增強它們生長和生存能力的輔助劑封包,然后培養該種胚,優選在無菌受控的條件下培養該種胚,以通過組織培養技術提供種間雜種(見例如m.monnier,"cultureofzygoticembryos,"frontiersofplanttissueculture,t.a.thorpe,ed.theinternationalassociationforplanttissueculture,universityofcalgary,alberta,canada,pp.277-280,1978;ramonandhanneman,2002,introgressionofresistancetolateblight(phytophthorainfestans)fromsolanumpinnatisectumintos.tuberosumusingembryorescueanddoublepollination.euphytica127(3):421-435)。可選擇地,可以對來自性狀供體的基因進行克隆,且通過如下面所述的轉化技術將克隆的基因引入受體植物中。能預見可以開發基因標記物用于監控后代植物中的性狀的存在。可以通過發現與性狀相關聯的基因組序列(如通過連鎖作圖研究)來進行所述標記物的開發。連鎖圖譜是示出其已知基因或基因標記物就重組頻率而言相對于彼此的位置的物種或實驗種群的基因圖譜,而不是已知基因或基因標記物沿每一條染色體的具體物理距離。連鎖圖譜是基于在同源染色體的交叉的過程中標記物之間重組頻率的圖譜。兩個基因標記物之間重組(分離)頻率越高,則假定它們相距越遠。相反,標記物之間的重組頻率越低,則它們之間的物理距離越小。可使用確認或假定的非編碼dna序列(如微衛星或那些生成限制性片段長度多態性(rflp)、aflp、snp、caps、rapd、est、ssr或issr標記物)作為標記物,且可共同鑒定一條染色體上示出與感興趣的表型性狀相連鎖的多個相鄰標記物以定義該性狀的定量性狀基因座(qtl)。技術人員應意識到這些技術。b.通過標記物輔助的育種的基因滲入在可用作性狀來源的供體加入物或供體品種是已知的,且可輔助選擇那些可能包含新型性狀的等位基因的后代植物的基因標記物是可用的情況下,使用下面的方法。提供包括期望性狀的(野生型)加入物或品種的植物,且在所述性狀供體品系的植物和來自根據本發明的基本純合的育種品系的植物之間進行雜交。通過進行標記物輔助選擇的步驟從雜種后代植物中選擇可能顯示出所述性狀的植物。至此,測試后代(其中所述后代可以為種子、種胚或植物的形式)的核酸(優選dna)是否存在與所述性狀有聯系的標記物,以便確認所述后代中是否潛在存在所述性狀的等位基因。標記物可以是微衛星、rflp、aflp、snp、caps、rapd、est、ssr或issr標記物,或技術人員可用的任何其它標記物。然后使用根據本發明的基本純合的受體育種品系作為回歸親本進行重復回交,且通過標記物輔助選擇來選擇可能包含上述性狀的等位基因的后代植物,以監控包含產生期望性狀的基因的基因組片段從性狀供體至受體育種品系的成功轉移,其中所述回交步驟用以保持或增加子代中受體育種品系的基因組背景或基因貢獻(geneticcontribution)。進行重復回交和選擇的過程,以便留下供體品系的不期望的性狀而使感興趣的基因轉移至受體育種品系中。可以通過選擇對回歸親本(基因組)特異性的(隨機)標記物來降低需要基本恢復回歸親本基因組的世代的數量。最后,通過選擇與期望等位基因聯系最緊密的重組事件,可限制基因累贅(geneticdrag)。然后,這些步驟提供了具有使農藝學期望性狀的額外基因并入其基因組中的馬鈴薯育種品系。最后,由這些回交和選擇步驟所得到的植物自交以提供其中并入額外期望性狀的純合育種品系。在由回交步驟所得到的所有后代植物中和/或優選在純合育種品系中,可以通過表型篩選(例如生物檢驗)確認期望性狀的存在。在性狀遺傳(顯性對隱性;單基因對多基因)的基因信息是已知的情況下,可以對回交和選擇步驟進行優化以實現性狀從供體至基本純合的受體育種品系的成功轉移。技術人員應意識到優化前面所述的轉移所需要的各種步驟。在包括期望性狀的加入物不能與來自根據本發明的基本純合的受體育種品系的植物雜交的情況下(即如果它們是不可雜交的物種),可使用上述的種胚挽救方法。可選擇地,可以對來自性狀供體的基因進行克隆,且通過如本文下面所述的轉化技術將克隆的基因引入受體植物中。可通過本發明的該方面產生的植物(其中進行使用已知的性狀供體進行標記物輔助育種)包括如下性狀:對晚疫病菌的抗性,在如vandervossen等.(plantj.36(6):867-882,2003)中所述的標記物ct88-ct64之間的solanumbulbocastanum染色體7上的基因座rpi上;對晚疫病菌的抗性,存在于solanumdemissum染色體11上的基因座r6-r11上,如在el-kharbotly等.1996;black等.,(1953);和huangs(2005)中所述;及進一步如在表10中示出的。因此,本發明涉及由兩個基本純合的親本育種品系之間雜交所得到的雜種植物,該兩個基本純合的親本育種品系中的至少一個已經在其基因組中并入且提供給f1雜種以下性狀:對來自solanumbulbocastanum或solanumdemissum的晚疫病菌的抗性;對來自solanumtuberosum的亞種andigenacpc1690或1673的g.rostochiensisro1的抗性;對來自solanumtuberosum亞種andigena的g.pallida的抗性;對來自solanumtuberosum登錄號78-3778的globoderarostochiensisro5-22和g.pallidapa2-d383的抗性;對g.pallidapa2和pa3vanwelkeaccessie????的抗性;對來自solanumtuberosum登錄號3704-76的g.pallidapa3的抗性;對來自solanumbulbocastanum的m.chitwoodi的抗性;對來自solanumfendlerii和solanumhougassii的m.chitwoodi/m.fallax的抗性;對來自solanumchacoense和solanumtarijense的m.hapla的抗性;對來自solanumtuberosumcvpentlandivory的pvx的抗性;以及對來自solanumtuberosumdw83-3121的pvs的抗性。c.已知基因進入二倍體、基本純合且自交親和的育種品系在可用作性狀來源的供體加入物或供體品種是已知的,且在該來源中產生所述性狀的基因序列是可用的情況下,可使用下面的方法用于將該基因并入受體育種中。可開發基于核酸檢測的測試,用于確定植物部分中的基因的存在,包括例如用于以序列特異性方式(例如pcr)擴增所述基因(的一部分)的擴增引物。這樣的測試可以用在如上所述的標記物輔助育種的方法中,作為基于連鎖圖譜研究檢測與所述形狀相關聯的標記物的替代方式。基因檢測的優點是可以以單一的檢驗明確確認基因的存在,因為基因序列可以提供具有100%可靠性的診斷標記物。因此,提供包括期望的額外性狀的(野生型)加入物或品種的植物,且在所述性狀供體品系的植物和來自基本純合的受體育種品系的植物之間進行雜交。通過在后代植物中進行確定賦予性狀的基因存在的步驟,從后代植物中選擇顯示出所述基因的植物。至此,通過使用用于序列特異性檢測的方法(如pcr檢測)測試后代(其中所述后代可以為種子、種胚或植物的形式)的核酸(優選dna)是否存在產生所述性狀的基因。然后使用基本純合的育種品系作為回歸親本進行重復回交,且通過使用如上所述的序列特異性檢測的方法從由此產生的后代中選擇包含所述基因的后代,以監控產生期望性狀的基因從性狀供體至基本純合的育種品系的成功轉移,其中所述回交步驟用以保持基因組背景或增加子代基因組中育種品系的基因貢獻。然后,這些步驟提供了具有受體品系關于農藝學期望性狀的所有特征,且進一步包括新的期望基因的馬鈴薯育種品系。最后,由這些回交和選擇步驟所得到的植物自交以提供其中以純合形式并入期望基因的純合育種品系。在由回交步驟所得到的所有后代植物中和/或優選在純合育種品系中,可以通過表型篩選(例如生物檢驗)確認期望性狀的存在。可選擇地,可以使用基因轉化的方法,其中從供體加入物或供體品種的染色體中分離基因,或其中產生合成的基因,且其中通過基因轉化技術將所述分離或合成的基因隨后引入根據本發明的受體品系中。根據本發明的一個方面,包括本文中定義的及下面表10的性狀中的一個或多個基因的核酸(優選dna)序列可以用于產生具有額外農藝學期望性狀的馬鈴薯育種品系的植物。在這個方面,本發明提供了本文定義的農藝學期望性狀的基因用于產生本文定義的基本純合的受體育種品系的用途,該用途包括將包括所述基因的核酸序列引入所述基本純合的受體育種品系的適當受體植物中。如上所述,所述核酸序列可以來源于適當的供體植物,且可以通過基因滲入(雜交)或通過轉化引入所述核酸序列。對于轉移包括農藝學期望性狀的期望基因的核酸序列的轉基因方法,可以通過本領域已知的方法從所述供體植物中分離,因此通過用于植物轉化的轉基因方法(例如通過載體的方式)將在配子中或在任何其它適當的轉移元件(如被所述核酸序列包裹顆粒進行的轟擊)中的分離的核酸序列轉移至受體植物。植物轉化通常包括具有將在植物細胞中行使功能的表達盒載體的構建。在本發明中,這樣的載體由包括期望基因的核酸序列組成,該載體可包括在調節元件(如啟動子)控制下的或與調節元件可操作地連接的基因。表達載體可包含這樣的一個或多個可操作地連接的基因/調節元件的組合,但前提是組合中包含的至少一個基因賦予農藝學期望性狀。所述載體可以是質粒的形式,且所述載體可以單獨使用或與其它質粒組合使用,以使用本領域已知的轉化方法(如農桿菌(agrobacterium)轉化系統)提供包含所述基因的轉基因植物。還以轉基因方法的形式,可以從供體植物中分離包括農藝學期望性狀的期望基因的核酸序列,該供體植物可以與種植的馬鈴薯(s.tuberosum)雜交。這樣的核酸序列被定名為同源基因(cisgene)(jacobsen&schouten,2008,pot.res.51:75-88)。同源基因經由本文所述的轉化技術引入馬鈴薯(定名為“同源轉基因(cisgenesis)”或“基因內途徑”)被認為是基因轉化技術,由此生成的產物的安全性與傳統育種或突變育種相似(rommens等.,2007,plantscience12:397-403;jacobsenandschouten,2008,pot.res.51:75-88;rommens,2010,biotechnologyinagricultureandforestry64,61-77)。現今,同源轉基因已經應用在馬鈴薯中(jacobsenandschouten,2008,pot.res.51:75-88;www.cisgenesis.com)。表達載體可包括可操作地連接至調節元件(如啟動子)的至少一個標記物基因,從而允許通過負選擇(通過抑制不包含可選擇的標記物基因的細胞的生長),或通過正選擇(通過篩選由標記物基因編碼的產物)來找回包含標記物的轉化的細胞。很多普遍使用的用于植物轉化的可選擇的標記物基因在本領域是已知的,且包括例如:編碼新陳代謝地除去選擇性化學試劑的毒性的酶的基因,該化學試劑可以是抗生素或除草劑;或編碼改變的目標的基因,該改變的目標對抑制劑不敏感。幾種正選擇方法在本領域是已知的,如甘露糖選擇。可選擇地,可以使用標記物-較少的轉化以獲得沒有提到的標記物的基因的植物,用于該過程的技術在本領域是已知的。一種用于將表達載體引入植物中的方法是基于農桿菌的天然轉化系統(見例如horsch等.,1985,science227:1229-1231)。a.tumefaciens和a.rhizogenes是基因轉化植物細胞的植物致病性土壤細菌。a.tumefaciens和a.rhizogenes的ti和ri質粒分別攜帶用于植物基因轉化的基因。將表達載體引入植物組織的方法包括直接感染或植物細胞與agrobacteriumtumefaciens共培養。在美國專利號5,591,616中提供了農桿菌載體系統的描述和用于農桿菌介導的基因轉移的方法。可以在gruber和crosby,1993(見gruber和crosby,1993.vectorsforplanttransformation.in:b.r.glick和j.e.thompson,編輯methodsinplantmolecularbiologyandbiotechnology,crcpress,bocaraton,第89-119頁中發現植物表達載體和報告基因和轉化方案的普通描述及農桿菌載體系統和用于農桿菌介導的基因轉移的方法的描述。例如miki等,1993(見miki等,1993.procedureforintroducingforeigndnaintoplants.in:methodsinplantmolecularbiologyandbiotechnology,ed.glickandthompson,crcpress,bocaraton,pages67-88)提供了培養植物組織的普通方法。用于分子克隆技術和適當的表達載體的適當的參考書是sambrook和russell的《分子克隆實驗指南(molecularcloning:alaboratorymanual)(第三版)》(冷泉港實驗室出版社,2001)。另一用于將表達載體引入植物中的方法是基于微粒介導的轉化(微粒轟擊),其中,dna被攜帶在微粒的表面。表達載體經生物槍裝置(biolisticdevice)被引入植物組織中,該生物槍裝置使微粒加速至足以穿透植物細胞壁和膜的300~600m/s的速度。另一用于將dna引入植物中的方法是經由目標細胞的超聲波處理(sonication)。可選擇地,脂質體或原生質球融合可用以將表達載體引入植物中。也可以使用經cacl2沉淀、聚乙烯醇或多聚鳥氨酸將dna直接攝取入原生質體,如可以是原生質體及整個細胞和組織的電穿孔。其它基于在大腸桿菌中發現的天然存在的f-因子的質粒的公知的技術(如使用bac)可例如被用于與bibac系統組合以產生轉基因植物,其中馬鈴薯基因組的一部分被引入細菌人工染色體(bac)中,細菌人工染色體(bac)即為用以在大腸桿菌(escherichiacoli)細胞中克隆dna片段(100-bp至300-kb插入大小;平均,150kb)的載體。馬鈴薯目標組織轉化之后,上述可選擇的標記物基因的表達允許使用本領域中現在公知的再生和選擇方法優選選擇被轉化的細胞、組織和/或植物。可以通過本發明的該方面產生的植物,其中被引入受體育種品系中的已知的馬鈴薯基因包括那些含在表10中示出的黃色果肉顏色、橙色果肉顏色、直鏈淀粉/支鏈淀粉、制片品質、低溫糖化、淀粉含量和炸薯條品質(frenchfryquality)、黑斑瘀傷、酚類抗氧化劑、丙烯酰胺水平、淀粉磷酸鹽含量和油炸品質、對晚疫病菌的抗性、對g.rostochiensisro1的抗性及對馬鈴薯病毒y(pvy)和馬鈴薯病毒x(pvx)的抗性的性狀,及由genbank登錄號hm013963.1、hm013964.1、hm013965.1、x58453.1、x70368.1、eu684230.1、x73477.1、aj133765、hq110081、hq197977、ay341425.1、ay864819.1、ay864820.1、fj810205.1、fj810206.1、fj810207.1、fj810208.1、fj810209.1、ay864821.1、gu980593.1、gu980594.1、gu980595.1、u22921.1、u22922.1、u22923.1、ay205302.1、eu310399.1、ck278037.1、ck271149.1、af143202.1、y09533.1、fj536325.1、fj536323.1、gu563971.1、gu563972.1、gu563973.1、fj536324.1、fj423045.1、fj423046.1、ay336128.1、dq122125.1、fj188415.1、ay196151.1、af195939.1、y15297.1和aj300266.1的基因賦予的性狀。在本文中使用的所有genbank登錄號指ncbi-genbankflatfilerelease183.0在2011年4月21日的基因序列數據庫(geneticsequencedatabank)。因此,本發明涉及由兩個基本純合的親本育種品系之間雜交得到的雜種植物,該兩個基本純合的親本育種品系中的至少一個已經在其基因組中并入且提供給f1雜種:β-胡蘿卜素羥化酶2(chy2)的基因、玉米黃質環氧化酶(zep)的基因、顆粒結合淀粉合成酶的馬鈴薯基因、β-呋喃果糖苷酶或馬鈴薯酸性轉化酶的基因、蔗糖磷酸合成酶的基因、假定轉化酶抑制劑前體(putativeinvertaseinhibitorprecursor)(inh1/inh2)的基因、塊莖多酚氧化酶ppo的基因、蔗糖合成酶的基因、綠原酸誘導因子(chlorogenateinducer)的基因、天冬酰胺合成酶的表達序列標簽est724115和est717227、磷酸化酶-l基因、r1蛋白或葡聚糖水二激酶(glucanwaterdikinase)的基因,這些基因均來自solanumtuberosum;s.demissum的r2和r2-樣蛋白基因,.hjertingii的r2晚疫病抗性蛋白(rpi-hjt1.3)的基因,abptrpi蛋白基因,s.venturii晚疫病抗性蛋白,s.bulbocastanum假定的疾病抗性蛋白rga2的基因,s.bulbocastanum晚疫病抗性蛋白rpi的基因,s.bulbocastanum晚疫病抗性蛋白rpi-blb2(rpi-blb2)的基因,s.bulbocastanum的nbs-lrr抗性蛋白(rpi-bt1)的基因,s.tuberosum線蟲抗性蛋白(gro1-4)的基因,s.tuberosum的疾病抗性蛋白gpa2的基因,s.tuberosum亞種andigena的adg2/2x(v-2)7a基因控制的對馬鈴薯y和a馬鈴薯y病毒屬(potyviruses)的抗性,和/或s.tuberosum亞種的抗性基因-樣、外顯子1~6、剪接變體c38和c19的andigenary-1基因。d.可用的轉基因植物的用途根據本發明,可以使用之前已經整合入馬鈴薯基因組中的轉基因序列。可以通過與攜帶期望性狀的基因的轉基因植物進行雜交來生成本發明的植物。可選擇地,可以經由轉化技術將轉基因直接引入育種品系的植物中。可以在本發明的該方面中使用的轉基因馬鈴薯植物包括:包含如在表10中示出的紅色花青素、藍色或紫色色素,塊莖皮的色素沉著,對晚疫病菌的抗性的那些轉基因的性狀;及由genbank登錄號ay289924.1、ay675561.1、ay675560.1、ay675558.1、ay675559.1、ay841129.1、af447489.1、ay849382.1和fj188415.1的基因所賦予的性狀。所有在本文中使用的genbank登錄號指ncbi-genbankflatfilerelease183.0在2011年4月21日的基因序列數據庫。因此,本發明涉及由兩個基本純合的親本育種品系之間雜交得到的雜種植物,該兩個基本純合的親本育種品系中的至少一個已經在其基因組中并入且提供給f1雜種:二氫黃酮醇4-還原酶基因、類黃酮3’,5’-羥化酶基因和/或花青素-1產生途徑的r2r3myb轉錄調節因子的基因,這些基因均來自s.tuberosuml.;s.demissum的晚疫病抗性蛋白(r1)基因,來自s.tuberosuml.的馬鈴薯晚疫病抗性蛋白r3a的基因,及s.bulbocastanumnbs-lrr抗性蛋白(rpi-bt1)的基因。為了制得純的或近交增加的品系,如上所述的均具有所述基因滲入或轉化的馬鈴薯育種品系的所有植物可以通過與回歸親本連續回交的步驟產生良種品系。因此,在本文所述的本發明的所有方面中,本發明還提供了優秀的或近交的馬鈴薯品系。包含額外期望性狀的本發明的基本純合的育種品系的植物是二倍體,且可以通過本領域已知的方法開發成四倍體植物。實例8包含額外期望性狀的本發明的基本純合的育種品系的植物是二倍體,且可以在雜交中用作親本以產生商品化的f1雜種(或品種)。包含額外期望性狀的的本發明的基本純合的育種品系的植物包含如在表10中所述性狀或基因中的至少一種、優選至少兩種或三種、更優選至少四種、仍然更優選至少五種。包含額外期望性狀的的本發明的基本純合的育種品系的植物優選包含選自表10中與塊莖性狀相關的基因中的至少一個基因,優選與選自表10中與處理或塊莖品質性狀相關的基因中的至少一個基因組合,優選進一步與選自表10中的與晚疫病菌抗性有關的基因中的至少一個基因組合,優選進一步與選自表10中的與線蟲抗性相關的基因中的至少一個基因組合,優選進一步與選自表10中的與病毒抗性相關的基因中的至少一個基因組合。可選擇地,根據本發明的基本純合的育種品系的植物包含至少兩個基因,該兩個基因中的一個選自表10中與塊莖性狀相關的基因,且另一個選自表10中與處理或塊莖品質性狀相關的基因,或選自表10中與晚疫病菌抗性相關的基因,或選自表10中與線蟲抗性相關的基因,或選自表10中與病毒抗性相關的基因。在又一可選擇的方式中,根據本發明的基本純合的育種品系的植物包含至少兩個基因,該兩個基因中的一個選自表10中與處理或塊莖品質性狀相關的基因,且另一個選自表10中與晚疫病菌抗性相關的基因,或選自表10中與線蟲抗性相關的基因,或選自表10中與病毒抗性相關的基因。在又一可選擇的方式中,根據本發明的基本純合的育種品系的植物包含至少兩個基因,該兩個基因中的一個選自表10中與晚疫病菌抗性相關的基因,且另一個選自表10中與線蟲抗性相關的基因,或選自表10中與病毒抗性相關的基因。在又一可選擇的方式中,根據本發明的基本純合的育種品系的植物包含至少兩個基因,該兩個基因中的一個選自表10中與線蟲抗性相關的基因,且另一個選自表10中與病毒抗性相關的基因。可選擇地,根據本發明的基本純合的育種品系的植物包含至少三個基因,該三個基因中的第一個選自表10中與塊莖性狀相關的基因,第二個選自表10中與處理或塊莖品質性狀相關的基因,且第三個選自表10中與晚疫病菌抗性相關的基因,或選自表10中與線蟲抗性相關的基因,或選自表10中與病毒抗性相關的基因。在又一可選擇的方式中,根據本發明的基本純合的育種品系的植物包含至少三個基因,該三個基因中的第一個選自表10中與處理或塊莖品質性狀相關的基因,第二個選自與晚疫病菌抗性相關的基因,且第三個選自表10中與線蟲抗性相關的基因,或選自表10中與病毒抗性相關的基因。在又一可選擇的方式中,根據本發明的基本純合的育種品系的植物包含至少三個基因,該三個基因中的第一個選自表10中與晚疫病菌抗性相關的基因,第二個選自表10中與線蟲抗性相關的基因,且第三個選自表10中與病毒抗性相關的基因。表10.適用于并入二倍體馬鈴薯育種品系的性狀。技術人員使用根據本發明的上述任何基本純化的親本育種品系均能夠創建多樣的可能的親本品系組合。因此,在雜交中使用這些親本品系中的兩個以產生f1馬鈴薯品種可得到同等多樣的可能f1雜種。通過篩選以下三種特征中任一種可容易地將本發明的雜種植物(f1雜種)與現有技術的馬鈴薯雜種區分開:(基因)一致性、低等位基因頻率,和提供有活力的加倍的(雙)單倍體。基因一致性根據本發明的雜種品系顯示出一致性,意味著在通過使兩個基本純合的育種品系(至少其中一個是根據本發明的純合的育種品系)雜交所產生的品系或品種內植物特征從植物至植物是一致的。相反,現有技術的雜種馬鈴薯品系沒有示出一致性。事實上,在現有技術的馬鈴薯品系中,相同品系的不同植物的特征是不同的。雖然生長增殖的傳統馬鈴薯品種也是基因一致的,但是當這樣傳統的品種由種子開始種植(真正的馬鈴薯種子,tps)時,植物不是基因一致的。因此,與真正的馬鈴薯種子相比,根據本發明各方面的馬鈴薯品種是基因一致的且獨特的。因此,可以通過測試植物相對于該品系的其它植物是否是一致的來識別根據本發明的雜種植物。低等位基因頻率在傳統四倍體馬鈴薯品種和育種克隆中超過80%的基因座是四基因的,意味著四個不同的等位基因存在于四倍體基因組的基因座上。3~5%是三基因的,且少于1%是二基因的。因此,存在顯著高頻率的三基因和四基因的基因座。本發明的二倍體馬鈴薯雜種可以進行染色體加倍,從而得到四倍體雜種。由此產生的四倍體雜種沒有三基因或四基因的基因座,且因此容易使用分析等位基因頻率的標記物分析與現有技術的四倍體相區分。有活力的加倍的(雙)單倍體本發明人已經從二倍體馬鈴薯品系生成四倍體品系和雜種。有一些關于從四倍體馬鈴薯生成雙單倍體和加倍的雙單倍體的文獻。結果是從來沒有從加倍的(雙)單倍體植物生成出有活力且能育的子代。這很可能是因為可頻繁出現在傳統馬鈴薯品種中的有害的隱性基因。由于有害等位基因已經在育種過程中被去除的事實,如本文所述的來自新型馬鈴薯品系的加倍(雙)單倍體是有活力的。因此,本發明各方面的雜種(f1)馬鈴薯品系容易與現有技術的二倍體品系相區分,顯現為有活力且能育的加倍的(雙)單倍體。表10的參考文獻1.bakker,e.,butterbach,p.,rouppevandervoort,j.,vandervossen,e.,vanvliet,j.,bakker,j.&goverse,a.(2003).geneticandphysicalmappingofhomologuesofthevirusresistancegenerx1andthecystnematoderesistancegenegpa2inpotato.theor.appl.genet.106:1524-15312.ballvora,a.,ercolano,m.r.,weiss,j.,meksem,k.,bormann,c.a.,oberhagemann,p.,salamini,f.&gebhardt,c.(2002).ther1geneforpotatoresistancetolateblight(phytophthorainfestans)belongstotheleucinezipper/nbs/lrrclassofplantresistancegenes.plantj.30:361-3713.bendahmane,a.,kanyuka,k.&baulcombe,d.c.(1999).therxgenefrompotatocontrolsseparatevirusresistanceandcelldeathresponses.plantcell11:781-7924.black,w.,mastenbroek,c.,mills,w.r.&peterson,l.c.(1953).aproposalforaninternationalnomenclatureofracesofphytophthorainfestansandofgenescontrollingimmunityinsolanumdemissumderivatives.euphytica2:173-1795.bonierbale,m.w.,plaisted,r.l.&tanksley,s.d.(1988).rflpmapsbasedonacommonsetofclonesrevealmodesofchromosomalevolutioninpotatoandtomato.genetics120:1095-11036.bradshaw,j.e.,meyer,r.c.,milbourne,d.,mcnicol,j.w.,phillips,m.s.&waugh,r.(1998).identificationofaflpandssrmarkersassociatedwithquantitativeresistancetogloboderapallida(stone)intetraploidpotato(solanumtuberosumsubsp.tuberosum)withaviewtomarker-assistedselection.theor.appl.genet.97,202-2107.brown,c.r.,yang,c-p.,mojtahedi,h.,santo,g.s.&masuelli,r.(1996).rflpanalysisofresistancetocolumbiaroot-knotnematodederivedfromsolanumbulbocastanuminabc2population.theor.appl.genet.92:572-5768.brown,c.r.,mojtahedi,h.,&santo,g.s.(2003).characteristicsofresistancetocolumbiaroot-knotnematodeintrogressedfromseveralmexicanandnorthamericanwildpotatospecies.actahort.619:117-1259.brown,c.r.,kim,t.s.,ganga,z.,haynes,k.,dejong,d.,jahn,m.,paran,i.&dejong,w.(2006).segregationoftotalcarotenoidinhighlevelpotatogermplasmanditsrelationshiptobeta-carotenehydroxylasepolymorphism.amer.j.potatores.83:365-37210.brummell,d.a.,chen,r.k.y.,harris,j.c.,zhang,h.,hamiaux,c.,kralicek,a.v.&mckenzie,m.j.(2011).inductionofvacuolarinvertaseinhibitormrnainpotatotuberscontributestocold-inducedsweeteningresistanceandincludessplicedhybridmrnavariants.j.exper.bot.,advancedaccessmarch10,2011pg1-16(doi:10.1093/jxb/err043)11.champouret,n.(2010).functionalgenomicsofphytophthorainfestanseffectorsandsolanumresistancegenes.phdthesis.wageningenuniversity12.dejong,h.(1987).inheritanceofpigmentedtuberfleshincultivateddiploidpotatoes.amer.potatoj.64:337-34313.dejong,w.,forsyth.a.,leister.d.,gebhardt,c.&baulcombe,d.c.(1997).apotatohypersensitiveresistancegeneagainstpotatovirusxmapstoaresistancegeneclusteronchromosome5.theor.appl.genet.95:246-25214.dejong,w.s.,dejong,d.m.,dejong,h.,kalazich,j.&bodism.(2003).analleleofdihydroflavonol4-reductaseassociatedwiththeabilitytoproduceredanthocyaninpigmentsinpotato(solanumtuberosuml.).theor.appl.genet.107:1375–138315.draaistra,j.(2006).geneticanalysisofroot-knotnematoderesistanceinpotato.phdthesiswageningenuniversity,thenetherlands.http://edpot.wur.nl/4390216.el-kharbotly,a.,leonards-schippers,c.,huigen,d.j.,jacobsen,e.,pereira,a.,stiekema,w.j.,salamini,f.&gebhardt,c.(1994).segregationanalysisandrflpmappingofther1andr3allelesconferringracespecificresistancestophytophthorainfestansinprogeniesofdihaploidpotatoparents.mol.gen.genet.242:749-754.17.el-kharbotly,a.,paloino-sánchez,c.,salamini,f.,jacobsen,e.&gebhardt,c.(1996).r6andr7allelesofpotatoconferringrace-specificresistancetophytophthorainfestans(mont.)debaryindentifiedgeneticlociclusteringwithther3locusonchromosomexi.theor.appl.genet.92:880-884.18.ewing,e.e.,i,simko,c.d.smart,m.w.bonierbale,e.s.g.mizubuti,g.d.may&w.e.fry(2000).geneticmappingfromfieldtestsofqualitativeandquantitativeresistancetophytophthorainfestansinapopulationderivedfromsolanumtuberosumandsolanumberthaultii.mol.br.6:25-36.19.fan,c.y.,yin,j.-m.,wang,b.,zhang,y.f.&yang,q.,(2010).molecularcloningandexpressionanalysisofafthomologousgenefromsolanumtuberosum.agricult.sci.china9:1133-113920.foster,s.j.,park,t.h.,pel,m.,brigneti,g.,j.,jagger,l.,vandervossen,e.&jones,j.d.g.(2009).rpi-vnt1.1,atm-22homologfromsolanumventurii,confersresistancetopotatolateblight.mol.pl.microb.inter.22:589-60021.gebhardt,c.,mugniery,d.,ritter,e.,salamini,f.&bonnel,e.(1993).identificationofrflpmarkerscloselylinkedtotheh1geneconferringresistancetogloboderarostochiensisinpotato.theor.appl.genet.85:541-54422.j.h.,watanabe,k.n.,valkonen,j.p.t.,arihara,a.,plaisted,r.l.,pehu,e.,miller,l.&slack,s.a.(1997).mappingandmarker-assistedselectionforageneforextremeresistancetopotatovirusy.theor.appl.genet.94:192-197.23.huang,s.,vleeshouwers,v.g.a.a.,werij,j.s.,hutten,r.c.b.,vaneck,h.j.,visser,r.g.f.&jacobsen,e.(2004).ther3resistancetophytophthorainfestansinpotatoisconferredbytwocloselylinkedrgeneswithdistinctspecificities.mol.pl.micr.interact.17:428-43524.huang,s.(2005).discoveryandcharacterizationofthemajorlateblightresistancecomplexinpotato.thesis,wageningenuniversity,thenetherlands.25.huang,s.,vandervossen,e.a.,kuang,h.,vleeshouwers,v.g.,zhang,n.,borm,t.j.,vaneck,h.j.,baker,b.,jacobsen,e.&visser,r.g.(2005).comparativegenomicsenabledtheisolationofther3alateblightresistancegeneinpotato.plantj.42:251-26126.jung,c.s.,griffiths,h.m.,dejong,d.m.,cheng,s.,bodis,m.&dejong,w.s.(2005).thepotatoplocuscodesforflavonoid3’,5’-hydroxylase.theor.appl.genet.110:269-27527.jung,c.s.,griffiths,h.m.,dejong,d.m.,cheng,s.,bodis,m.,kim,t.s.&dejong,w.s.(2009).thepotatodeveloper(d)locusencodesanr2r3mybtranscriptionfactorthatregulatesexpressionofmultipleanthocyaninstructuralgenesintuberskin.theor.appl.genet.120:45-5728.kanyuka,k.,bendahmane,a.,rouppevandervoort,j.n.a.m.,vandervossen,e.a.g.&baulcombed.c.(1999).mappingofintra-locusduplicationsandintrogresseddna:aidstomap-basedcloningofgenesfromcomplexgenomesillustratedbyphysicalanalysisoftherxlocusintetraploidpotato.theor.appl.genet.98:679-689.29.kasai,k.,morikawa,y.,sorri,v.a.,valkonen,j.p.t.,gebhardt,c.&watanabe,k.n.(2000).developmentofscarmarkerstothepvyresistancegeneryadgbasedonacommonfeatureofplantdiseaseresistancegenes.genome43:1-8.30.kim,h.s.,jeon,j.h.,choi,k.h.,joung,y.h.,lee,b.i.&joung,h.(2000).regulationofstarchcontentsinpotato(solanumtuberosuml.)bymanipulationofsucrosesynthasegene.j.jap.soc.hort.sci.69:243-249.31.kreike,c.m.,dekoning,j.r.a.,vinke,j.h.,vanooijen,j.w.&stiekema,w.j.(1994).quantitativelyinheritedresistancetogloboderapallidaisdominatedbyonemajorlocusinsolanumspegazzinii.theor.appl.genet.88:764-76932.kuhl,j.c.,hanneman,r.e.&havey,m.j.(2001).characterizationandmappingofrpi1,alateblightresistancelocusfromdiploid(1ebn)mexicansolanumpinnatisectum.mol.gen.genet.265:977-98533.leonards-schippers,c.,gieffers,w.,r.,ritter,e.,knapp,s.j.,salamini,f.&gebhardt,c.(1994).quantitativeresistancetophytophthorainfestansinpotato:acasestudyforqtlmappinginanallogamousplantspecies.genetics137:67-77.34.li,l.,m.j.paulo,strahwald,j.,lubeck,j.,hofferbert,h.r.,tacke,e.,junghans,h.,wunder,j.,draffehn,a.,vaneeuwijk,f&gebhardt,c.(2008).naturaldnavariationatcandidatelociisassociatedwithpotatochipcolor,tuberstarchcontent,yieldandstarchyield.theor.appl.genet.116:1167-118135.lokossou,a.a.,park,t.h.,vanarkel,g.,arens,m.,ruyter-spira,c.,morales,j.,whisson,s.c.birch,p.r.j.,visser,r.g.f.,jacobsen,e.&vandervossen,e.a.g.(2009).exploitingknowledgeofr/avrgenestorapidlycloneanewlz-nbs-lrrfamilyoflateblightresistancegenesfrompotatolinkagegroupiv.mol.pl.micr.interact.22:630-64136.malcolmson,j.f.&black,w.(1966).newrgenesinsolanumdemissumlindl.andtheircomplementaryracesofphytophthorainfestans(mont.)debary.euphytica15:199-20337.marano,m.r.,malcuit,i.,dejong,w.&baulcombe,d.c.(2002).high-resolutiongeneticmapofnb,agenethatconfershypersensitiveresistancetopotatovirusxinsolanumtuberosum.theor.appl.genet.105:192-200.38.marczewski,w.,hennig,j.&gebhardt,c.(2002).thepotatovirussresistancegenensmapstopotatochromosomeviii.theor.appl.genet.105:564-567.39.moloney,c.,griffin,d.,jones,p.w.,bryan,g.j.,mclean,k.,bradshaw,j.e.&milbourne.d.(2010).developmentofdiagnosticmarkersforuseinbreedingpotatoesresistanttogloboderapallidapathotypepa2/3usinggermplasmderivedfromsolanumtuberosumssp.andigenacpc2802.theor.appl.genet.120:679-68940.oosumi,t.,rockhold,d.r.,maccree,m.m.,deahl,k.l.,mccue,k.f.&belknap,w.r.(2009).generpi-bt1fromsolanumbulbocastanumconfersresistancetolateblightintransgenicpotatoes.amer.j.potatores.86:456-46541.paal,j.,henselewski,h.,muth,j.,meksem,k.,menendez,c.m.,salamini,f.,ballvora,a.&gebhardt,c.(2004).molecularcloningofthepotatogro1-4geneconferringresistancetopathotypero1oftherootcystnematodegloboderarostochiensis,basedonacandidategeneapproach.plantj.38:285-29742.park,t.h.,foster,s.,brigneti,g.&jones,j.d.g.(2009).twodistinctpotatolateblightresistancegenesfromsolanumberthaultiiarelocatedonchromosome10.euphytica165:269-27843.pel,m.a.,foster,s.j.,park,t.h.,rietman,h.,vanarkel,g.,jones,j.d.,vaneck,h.j.,jacobsen,e.,visser,r.g.&vandervossen,e.a.g.(2009).mappingandcloningoflateblightresistancegenesfromsolanumventuriiusinganinterspecificcandidategeneapproach.mol.plantmicrob.interact.22:601-61544.pineda,o.,bonierbale,m.w.,plaisted,r.l.,brodie,b.b.&tanksley,s.d.(1993).identificationofrflpmarkerslinkedtotheh1geneconferringresistancetothepotatocystnematodegloboderarostochiensis.genome36:152-15645.rauscher,g.m.,smart,c.d.,simko,i.,bonierbale,m.,mayton,h.,greenland,a.&fry,w.e.(2006).characterizationandmappingofrpi-ber,anovelpotatolateblightresistancegenefromsolanumberthaultii.theor.appl.genet.112:674-687.46.ritte,g.,lloyd,j.r.,eckermann,n.,rottmann,a.,kossmann,j&steup,m.(2002).thestarch-relatedr1proteinisanα-glucan,waterdikinase.proc.natl.acad.sci.usa99:7166-7171.47.ritter,e.,debener,t.,barone,a.,salamini,f.&gebhardt,c.(1991).rflpmappingonpotatochromosomesoftwogenescontrollingextremeresistancetopotatovirusx(pvx).mol.gen.genet.227:81-8548.rommens,c.m.,ye,j.,richael,c.&swords,k.(2006).improvingpotatostorageandprocessingcharacteristicsthroughall-nativednatransformation.j.agric.foodchem.54:9882-988749.rommens,c.m.,richael,c.m.,yan,h.,navarre,d.a.,ye,j.,krucker,m.&swords,k.(2008a).engineerednativepathwaysforhighkaempferolandcaffeoylquinateproductioninpotato.plantj.6:870-88650.rommens,c.m.,yan,h.,swords,k.,richael,c.&ye,j.(2008b).low-acrylamidefrenchfriesandpotatochips.plantbiotech.j.6:843-853.51.rouppevandervoort,j.,lindeman,w.,folkertsma,r.,hutten,r.,overmars,h.,vandervossen,e.,jacobsen,e.&bakker,j.(1998).aqtlforbroad-spectrumresistancetocystnematodespecies(globoderaspp.)mapstoaresistancegeneclusterinpotato.theor.appl.genet.96:654-66152.rouppevandervoort,j.n.a.m.,janssen,g.j.w.,overmars,h.,vanzandvoort,p.m.,vannorel,a.,scholten,o.e.,janssen,.r,&bakker,j.(1999).developmentofapcr-basedselectionassayforroot-knotnematoderesistance(rmc1)byacomparativeanalysisofthesolanumbulbocastanumands.tuberosumgenome.euphytica106:187-19553.rouppevandervoort,j.,vandervossen,e.,bakker,e.,overmars,h.,vanzandvoort,p.,hutten,r.,klein-lankhorst,r.&bakker,j.(2000).twoadditiveqtlsconferringbroad-spectrumresistanceinpotatotogloboderapallidaarelocalizedonresistancegeneclusters.theor.appl.genet.101:1122-113054.salanoubat,m.&belliard,g.(1987).molecularcloningandsequencingofsucrosesynthasecdnafrompotato(solanumtuberosuml.):preliminarycharacterizationofsucrosesynthasemrnadistribution.gene60:47-5655.sattarzadeh,a.,achenbach,u.,lübeck,j.,strahwald,j.,tacke,e.,hofferbert,h.-r.,rothsteyn,t.&gebhardt,c.,(2006).singlenucleotidepolymorphism(snp)genotypingasbasisfordevelopingapcr-basedmarkerhighlydiagnosticforpotatovarietieswithhighresistancetogloboderapallidapathotypepa2/3.molec.breed.4:301-31256.song,j.,bradeen,j.m.,naess,s.k.,raasch,j.a.,wielgus,s.m.,haberlach,g.t.,liu,j.,kuang,h.,austin-phillips,s.,buell,r.,helgeson,j.p.&jiang,j.(2003).generbclonedfromsolanumbulbocastanumconfersbroadspectrumresistancetopotatolateblight.proc.natl.acad.sci.usa100:9128-913357.song,y.-s.,hepting,l.,schweizer,g.,hartl,l.,wenzel,g.&schwarzfischer,a.(2005).mappingofextremeresistancetopvy(rysto)onchromosomexiiusinganther-culture-derivedprimarydihaploidpotatolines.theor.appl.genet.111:879-887.58.song,y.-s.&schwarzfischer,a.(2008).developmentofstsmarkersforselectionofextremeresistance(rysto)topvyandmaternalpedigreeanalysisofextremelyresistantcultivars.amer.j.potatores.85:159-17059.sorri,v.a.,watanabe,k.n.&valkonen,j.p.t.(1999).predictedkinase-3amotifofaresistancegeneanalogueasauniquemarkerforvirusresistance.theor.appl.genet.99:164-17060.tan,m.y.a.,hutten,r.c.b,celis,c.,park,t.h.,niks,r.e.,visser,r.g.f.&vaneck,h.j.(2008).therpi-mcd1locusfromsolanummicrodontuminvolvedinresistancetophytophthorainfestans,causingadelayininfection,mapsonpotatochromosome4inaclusterofnbslrrgenes.mol.pl.micr.interact.21:909-91861.tan,m.y.a.,park,t.h.,alles,r.,hutten,r.c.b.,visser,r.g.f.&vaneck,h.j.(2009).gpaxitarloriginatingfromsolanumtarijenseisamajorresistancelocustogloboderapallidaandislocalisedonchromosome11ofpotato.theor.appl.genet.119:1477-148762.tan,m.y.a.,hutten,r.c.b,visser,r.g.f.&vaneck,h.j.(2010).theeffectofpyramidingphytophthorainfestansresistancegenesr(pi-mcd1)andr(pi-ber)inpotato.theor.appl.genet.121:117-12563.thygesen,p.w.,dry,i.b.&robinson,s.p.(1995).polyphenoloxidaseinpotato.amultigenefamilythatexhibitsdifferentialexpressionpatterns.pl.physiol.109:525-53164.vanderleij,f.r.,visser,r.g.,ponstein,a.s.,jacobsen,e.&feenstra,w.j.(1991).sequenceofthestructuralgeneforgranule-boundstarchsynthaseofpotato(solanumtuberosuml.)andevidenceforasinglepointdeletionintheamfallele.mol.gen.genet.228:240-24865.vandervossen,e.a.g.,rouppevandervoort,j.n.a.m.,kanyuka,k.,bendahmane,a.,sandbrink,h.,baulcombe,d.c.,bakker,j.,stiekema,w.j.&klain-lankhorsty,r.m.(2000).homologuesofasingleresistance-geneclusterinpotatoconferresistancetodistinctpathogens:avirusandanematode.plantj.23:567-57666.vandervossen,e.,sikkema,a.,hekkertbt,b.l.,gros,j.,stevens,p.,muskens,m.,wouters,d.,pereira,a.,stiekema,w.&allefs,s.(2003).anancientrgenefromthewildpotatospeciessolanumbulbocastanumconfersbroad-resistancetophytophthorainfestansincultivatedpotatoandtomato.plantj.36:867-88267.vaneck,h.j.,jacobs,j.m.,vandenberg,p.m.m.m.,stiekema,w.j.&jacobsen,e.(1994).theinheritanceofanthocyaninpigmentationinpotato(solanumtuberosum1.)andmappingoftuberskincolourlociusingrflps.heredity73:410-42168.vidal,s.,cabrera,h.,andersson,r.a.,fredriksson,a.&valkonen,j.p.t.(2002).potatogeney-1isanngenehomologthatconferscelldeathuponinfectionwithpotatovirusy.mol.pl.-micr.interact.15:717-72769.wolters,a.-m.a.,uitdewilligen,j.g.a.m.l.,kloosterman,b.a.,hutten,r.c.b.,visser,r.g.f.&vaneck,h.j.(2010).identificationofallelesofcarotenoidpathwaygenesimportantforzeaxanthinaccumulationinpotatotubers.pl.molec.biol.73:659–671.70.zhangy.,jung,c.s.&dejong,w.s.(2009).geneticanalysisofpigmentedtuberfleshinpotato.theor.appl.genet.119:143–150.71.zrenner,r.,salanoubat,m.,willmitzer,l.&sonnewald,u.(1995).evidenceofthecrucialroleofsucrosesynthaseforsinkstrengthusingtransgenicpotatoplants(solanumtuberosuml.).plantj.7:97-107.當前第1頁12
相關技術
網友詢問留言
已有0條留言
- 還沒有人留言評論。精彩留言會獲得點贊!
1