氯化兩面針堿在制備抗骨質疏松和骨丟失疾病中的應用的制作方法
本發明涉及藥物,具體涉及制備新適應證的藥物,尤其涉及氯化兩面針堿(Nitidine Chloride)在制備抗骨質疏松癥和骨丟失藥物中的應用。。
背景技術:
骨質疏松癥是多種原因引起的一組骨病,骨組織有正常的鈣化,鈣鹽與基質呈正常比例,以單位體積內骨組織量減少為特點的代謝性骨病變。在多數骨質疏松中,骨組織的減少主要由于骨質吸收增多所致。以骨骼疼痛、易于骨折為特征。
破骨細胞(osteoclast)是骨組織成分的一種,行使骨吸收的功能。破骨細胞起源于骨髓造血干細胞/單核細胞系統,是一種高度分化的多核細胞,其分化主要依賴于 RANKL 的刺激。 RANKL 屬于腫瘤壞死因子超家族成員,由成骨細胞,軟骨細胞與骨細胞分泌而來。在骨微環境中,RANKL 與前破骨細胞表面受體 RANK 結合,募集 RANK 信號傳導的轉接蛋白 TRAF6,啟動下游信號通路包括 NF-κB,MAPK,NFAT,AKT 以及鈣離子通道,促進破骨細胞分化成熟。其中 NF-κB 是破骨細胞分化成熟的關鍵信號,而 NFAT 介導細胞融合和破骨細胞分化,是破骨細胞終端分化的轉錄因子。同時,RANKL 還需要免疫受體酪氨酸活化基序(ITAM)介導的信號通路(其中包括破骨細胞相關受體(OSCAR),髓樣細胞觸發受體-2(TREM2),Fc受體共同γ鏈(FcRγ),以及DNAX活化蛋白12(DAP12)),來共同激活 NFAT,致使破骨細胞分化,發揮細胞功能。
當前,治療骨丟失疾病的方法多采用以下幾種:一、骨吸收抑制劑治療,例如雙磷酸鹽;二、激素替代療法。但是,研究發現應用激素替代療法,女性患乳腺癌的風險增大,并且抑制骨吸收的磷酸鹽制劑也在臨床應用中出現了較大的不良反應,這極大限制了臨床推廣。因此,尋找可以防治骨丟失疾病的藥物一直是人們夢寐以求的愿望。近幾年,中草藥的治療骨丟失疾病的研究逐漸開展起來。由于中草藥治療效果明顯且安全性較好,目前,已經發現部分中草藥單體具有抑制破骨細胞的作用,例如芒果苷,柚皮苷等。兩面針為蕓香科植物兩面針 Zanthorμlum nitidum(Roxb.)DC. 的干燥根,性味苦、辛,具有跌打損傷、活血化瘀等功效,是廣西特有中草藥。氯化兩面針堿是兩面針主要活性成分之一。既往的研究主要集中在抗腫瘤以及抗炎的治療。目前對破骨細胞形成的作用尚未有報道。
申請人研究氯化兩面針堿對骨質疏松癥的調控作用,通過TARcP 染色發現氯化兩面針堿呈劑量依賴性抑制 RANKL 誘導的破骨細胞形成與骨吸收作用; 同時還發現 1 μM 氯化兩面針堿也能夠干預人外周血單個核細胞體外誘導破骨細胞成熟分化。從作用機制上顯示,氯化兩面針堿是通過抑制 RANKL 激活的 NF-κB 與 NFATc1 信號通路來發揮作用,并下調了破骨細胞成熟分化標志基因(cathepsin K, D2, calcitonin receptor, NFATc1, TRAcP, OSCAR, FcRγ, TREM2, DAP12)的表達。再通過建立去卵巢小鼠的骨質疏松癥模型表明氯化兩面針堿可以保護卵巢切除導致的骨丟失。從 HE 染色及組織病理學結果分析,與去卵巢小鼠組相比,氯化兩面針堿治療組明顯降低了骨組織中破骨細胞的數量。
基于上述藥理作用,可擬開發氯化兩面針堿為制備治療骨質疏松癥和骨丟失的藥物。
技術實現要素:
本發明所要解決的技術問題在于研究氯化兩面針堿對破骨細胞的調控作用,設計在抗骨質疏松癥藥物中的應用。
本發明提供了氯化兩面針堿在制備康骨質疏松癥藥物中的應用。
申請人首次研究發現氯化兩面針堿劑量依賴性抑制破骨細胞形成和骨吸收功能;發現氯化兩面針堿通過干預 RANKL 激活的 NF-κB 和 NFAT 信號通路發揮其抑制破骨細胞的作用。為了進一步證明氯化兩面針堿對破骨細胞的作用,建立了小鼠去卵巢骨質疏松模型。結果表明,氯化兩面針堿給藥組能夠防治去卵巢誘發的骨丟失。綜上所述,氯化兩面針堿通過下調 RANKL 激活的 NF-κB 和 NFAT 來干預破骨細胞分化功能。
附圖說明
圖1兩面針堿對 RANKL 誘導的破骨細胞形成以及破骨細胞增殖、凋亡的影響
(A:兩面針堿化學結構。B: BMMs 在含有 100 ng/mL 的 M-CSF,100 ng/mL 的RANKL 以及不同濃度的兩面針堿的細胞分化液中培養 5 天,然后進行 TRAcP 染色。光學顯微鏡下觀察兩面針堿呈劑量依賴性的抑制 RANKL 誘導的破骨細胞形成。 C: 對 TRAcP 染色陽性多核細胞進行計數,對于細胞核 ≥3 歸為破骨細胞;
D:利用 Annexin V 凋亡試劑盒檢測不同濃度兩面針堿對破骨細胞的細胞凋亡的影響;E:利用 MTS 考察兩面針堿對 BMMs 孵育 48 小時后,藥物對細胞增殖的影響。*p<0.05, **p<0.005 , ***p<0.001)
圖2兩面針堿對 RANKL 誘導破骨細胞分泌的特異性基因的影響
(BMMs 在含有 100 ng/mL 的 M-CSF,100 ng/mL 的RANKL 以及不同濃度的兩面針堿(0,0.125,0.25,0.5 以及 1 μM)的細胞分化液中培養 7 天。利用 RT-PCR 檢測相關基因的 mRNA 表達水平。結果用 Image J 分析各條帶的灰度值,計算目的基因灰度值與 GAPDH 灰度值的比值,即為各自 mRNA 的相對表達水平,分析各組之間的差異)
圖3兩面針堿對骨吸收功能的影響
(經 RANKL 誘導 BMMs 分化后的破骨樣細胞,按照相同的數目接種于骨板上,并加入含有 RANKL 以及不同濃度的兩面針堿的完全培養基進行孵育 48 h; 對照組只含有 RANKL 的完全培養基。A: 通過 SEM 觀察各組的骨吸收面積(黑色箭頭標示)B:對骨吸收面積進行定量分析。*p<0.05, **p<0.005 , ***p<0.001)
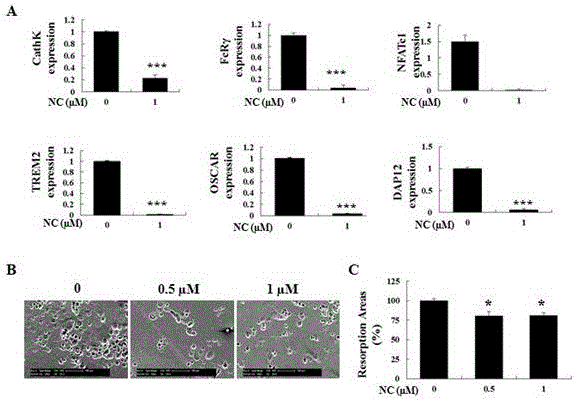
圖4兩面針堿對 RANKL 誘導的人外周血提取單核細胞相關基因的表達以及骨吸收功能作用
(A: 通過定量 PCR 比較,與只含有 RANKL 的完全培養基處理的陽性對照組比較, 1 μM 兩面針堿藥物處理組對 RANKL 誘導的人外周血提取單核細胞相關基因表達情況的影響 ;每個目的基因與相應的 GAPDH,進行比較循環閾值對目的基因進行計算;B:RANKL 誘導的人外周血提取單核細胞分化后的破骨樣細胞,按照相同的數目種于骨板上,并加入含有 RANKL 以及不同濃度的兩面針堿進行孵育 48 h;對照組只含有 RANKL 的完全培養基。通過 SEM 觀察各組的骨吸收面積;C:對骨吸收面積進行定量分析。*p<0.05, **p<0.005 , ***p<0.001)
圖5兩面針對 RANKL 引起的 IκBα 降解的影響
(采用穩定轉染 3κB-Luc-SV40 熒光素酶報告基因的 Raw 264.7,檢測硫酸小檗堿對 RANKL 活化 NF-κB 的影響。A: RAW 264.7 細胞分為3組, 陰性對照組含有完全培養基; 陽性對照組含有 100 ng/mL RANKL 完全培養基; 給藥組含有 100 ng/mL RANKL 與不同濃度的兩面針堿(0.5uM 和 1uM)的完全培養基。將各組數據進行比較,分析各組中各組中NF-κB 蛋白的表達情況。B:兩面針堿對BMMs進行預孵育 1h,然后按照時間點加入100ng/ml的RANKL 進行刺激。將 β-actin 最為內參,Western-blot 定量分析 IκBα 的表達。*p<0.05, **p<0.005 , ***p<0.001)
圖6兩面針堿對 RANKL 活化的 NFATc1 的作用
(采用穩定轉染 pNFATc1-TA-Luc 熒光素酶報告基因的 Raw 264.7,檢測硫酸小檗堿對 RANKL 活化 NFATc 的影響。A:RAW 264.7 細胞分為3組,陰性對照組-含有完全培養基;陽性對照組-含有 100 ng/ml 的RANKL的完全培養基; 給藥組-含有 100ng/ml 的RANKL和不同濃度的兩面針堿(0.5uM 和 1uM)的完全培養基。將各組數據進行比較,分析各組中 NFATc 蛋白的表達情況。 B:BMMs 在含有 100 ng/mL RANKL 與不同濃度的兩面針堿的完全培養基孵育 7 天;將 β-actin 最為內參,Western-blot 定量分析 NFATc1 與 d2 的表達。C:BMMs 預孵育兩面針堿(1uM) 1 h 后,按照時間點加入 100 ng/mL RANKL 與兩面針堿共孵育。將 β-actin 最為內參,Western-blot 定量分析 NFATc1 與 d2 的表達。*p<0.05, **p<0.005 , ***p<0.001)
圖7兩面針堿對去卵巢小鼠骨質疏松(OVX)引起的骨丟失的影響
(A:通過 Micro-CT 掃描形態測定分析小鼠脛骨近端干骺端骨小梁微結構的分析。 B: 脛骨標本分為 sham 組-假手術組,給藥組(兩面針堿 3 mg/kg 以及 6 mg/kg 腹腔注射 OVX 模型小鼠)以及雌激素組(50 ng/kg 腹腔注射 OVX 模型小鼠)。通過各組數據進行比較,分析各組小鼠骨體積分數(BV/TV)、骨小梁數量(Tb.N)、骨小梁間距(Tb.Sp)、骨小梁厚度(Tb/Th);實驗數據用均數±標準差表示;C:利用 HE 和 TRAcP 染色對骨小梁進行骨組織病理學檢測,分析各組骨小梁中骨吸收參數,例如破骨細胞表面積(OcS/BS), 破骨細胞數量 N.Oc/BS(n=3)。*p<0.05, **p<0.005 , ***p<0.001)。
具體實施方式
以下實施例用于說明本發明,但不用來限制本發明的范圍,若未特別說明,所用的技術手段為本領域技術人員所熟知的技術手段。
實施例1
一、材料與方法
試劑
氯化兩面針堿購自中國藥品生物制品檢定所(北京,中國)。RAW 264.7購自American Type Culture Collection(馬里蘭州,美國)。鼠源 MCSF 購自 R&D Systems (明尼阿波利斯,美國)。ɑ-MEM 購自 Thermo Fisher,胎牛血清(FBS)購自 TRACE (悉尼,澳大利亞)。重組 GST-rRANKL為本實驗室純化。人外周血單個核細胞誘導所需重組人源 M-CSF 購自 Chemicon (加尼福利亞洲,美國),胎牛血清,青鏈霉素和 L-谷氨酰胺購自 Invitrogen (加尼福利亞洲,美國)。 IκBα 和 NFATc1 抗體購自 Santa Cruz Biotechnology Com. (加尼福利亞洲,美國)。 V-ATPase d2 抗體來自本實驗室合成純化。
體外骨髓巨噬細胞的分離、培養和誘導分化
骨髓巨噬細胞由C57BL/6J小鼠的骨髓中分離獲取。實驗方法已經獲得西澳大學動物倫理委員會批準 (approval RA/3/100/1244)。骨髓巨噬細胞用含10% 胎牛血清,5 μg/mL青霉素,50 U/mL鏈霉素,2 mM L-谷氨酰胺以及10 ng/mL MCSF 的 ɑ-MEM 完全培養基,于 37℃ 含 5% CO2 培養箱孵育。
細胞長滿后,將BMMs(8×103)接種于96孔板,分為陰性對照組,陽性對照組,給藥組。其中陰性對照組不加 RANKL, 陽性對照組只加RANKL,給藥組加 RANKL 以及含不同濃度的氯化兩面針堿。每 2 天換液 1 次。7 天后,4% 的多聚甲醛固定細胞 20 分鐘,加入 TRAcP 染色液進行染色 1小時。光學顯微鏡下觀察 TRAcP 染色陽性多核細胞中細胞核 ≥3 個的細胞定義為破骨細胞,并進行計數。
)體外人外周血單個核細胞分離、培養和誘導分化
從澳大利亞紅十字會(阿德萊德,南澳大利亞)健康志愿者中采血。用預熱的HANK平衡液沖洗全血中的白膜層,并從聚蔗糖柱分理出人外周血單核細胞。實驗方法已經獲得西澳大學動物倫理委員會批準。用含有10% 胎牛血清,5 μg/mL青霉素,50 U/mL鏈霉素,2mM L-谷氨酰胺以及25 ng/mL MCSF 的 ɑ-MEM 完全培養液進行培養。并將細胞接種于12孔板用于mRNA檢測,以及含有骨板的 96 孔板用于骨吸收功能的檢測。次日,加入RANKL刺激分化并加入不同濃度的氯化兩面針堿。對照組加入 0.01% DMSO 進行培養。細胞孵育 7 天,每隔 2 天換 1 次液。
法測定氯化兩面針堿對破骨細胞前體增殖的影響
BMMs(6×103)接種于96孔板中培養過夜。更換培養基(含有不同濃度的氯化兩面針堿0.1,0.5,1, 5, 10 μM)孵育48小時。然后每孔加入 MTS 孵育2小時,最后進行酶標儀檢測。
雙染檢測凋亡
選用生長狀態良好的RAW 264.7細胞(1×106)接種于6孔板中培養過夜。更換培養基(含有不同濃度的氯化兩面針堿)進行孵育24小時后,用胰酶消化,收集細胞,用 PBS 沖洗1次,將細胞用 1× Binding緩沖液重懸(1×107 /mL),加入Annexin V-PI液染色。流式細胞儀進行分析,計算凋亡細胞的百分率。
反轉錄-聚合酶鏈反應
BMMs(1×105)接種于 6 孔板中培養過夜。更換培養基(含有不同濃度的氯化兩面針堿 0.5 和 1 μM)孵育 7 天。用 Trizol 裂解細胞并提取總 RNA。用含有oligo-dT 引物的反轉錄酶對 RNA 逆轉錄成 cDNA。然后加入不同基因的引物對 cDNA 進行聚合酶鏈反應擴增。18sRNA 作為內參。以下是不同基因引物序列:
18sRNA(Forward:ACCATAAACGATGCCGACT;Reverse:TGTCAATCCTGTCCGTGTC),
Calcitonin Receptor(Forward:TGGTTGAGGTTGTGCCCA;Reverse:CTCGTGGGTTTGCCTCATC),
cathepsin K (Forward:GGGAGAAAAACCTGAAGC;Reverse:ATTCTGGGGACTCAGAGC),
V-ATPase d2 (D2) (Forward:GGATCCGAATTCATGCTTGAGACTGCAGAG;Reverse:GGTCTAGATTATAAAATTGGAATGTAGCTC),
NFATc1 (mouse NFATc1) (Forward:CAACGCCCTGACCACCGATAG; Reverse:GGCTGCCTTCCGTCTCATAGT),
TRAcP (Forward:TGTGGCCATCTTTATGCT;Reverse:GTCATTTCTTTGGGGCTT)。
熒光定量PCR
將人外周血單個核細胞種于 12 孔板中并誘導分化 7 天后,Trizol 裂解細胞提取總 RNA。含有 oligo-dT 引物的反轉錄酶對 RNA 逆轉錄成 cDNA。將 cDNA 用 SYBR GREEN qPCR SuperMix-UDG 試劑在 Rotor-Gene 3000 擴增出目的基因 hARP,cathepsin K,NFATc1,OSCAR,FcRγ,TREM2,DAP12。所有樣本設三個副孔。擴增完成后,通過 Rotor-Gene Series 1.7 軟件按照 ddCT 法對目的基因變化進行相對變量。
熒光素酶報告基因
通過 2-二乙氨乙基葡聚糖將 pNFATc1-TA-Luc 穩定轉染 RAW 264.7。轉染的 RAW 264.7 細胞接種于 24 孔板過夜后,預孵育氯化兩面針堿 1 小時,加入100ng/mL RANKL刺激24小時。加入熒光素酶裂解緩沖液,移至白板,加入熒光素酶報告基因底物,熒光酶標儀檢測。
κB熒光素酶報告基因
通過 2-二乙氨乙基葡聚糖將 3κB-Luc-SV40 穩定轉染 RAW264.7。轉染的RAW 264.7 細胞接種于 24 孔板過夜后,預孵育氯化兩面針堿 1 小時,加入100ng/mL RANKL刺激8小時。加入熒光素酶裂解緩沖液,移至白板,加入熒光素酶報告基因底物,熒光酶標儀檢測。
骨吸收檢測
將 BMMs 接種于 6 孔板內,每孔加入含有 100ng/mL RANKL 和 10ng/mL mMCSF的完全培養基孵育3天。當破骨細胞形成,加入胰酶將細胞分離出來,并按照 1×103/well濃度種有 0.75mm 厚度骨板的96孔板內孵育過夜。加入氯化兩面針堿孵育48小時。其中一部分骨片用 4% 多聚甲醛固定,加入 TRAcP 染色液進行染色 1小時。光學顯微鏡下觀察 TRAcP 染色陽性多核細胞中細胞核 ≥3 個的細胞定義為破骨細胞,并進行計數。同時清洗另一部分骨片,晾干后,用 Philips XL30 掃描電鏡檢測骨吸收陷窩,并用Scion Image software計算骨吸收面積百分比。
牙本質片吸收陷窩形成的檢測
人外周血單個核細胞接種在置有牙本質片的96孔板內,加入 RANKL 刺激10天。加入胰酶消化細胞,并清洗牙本質片。晾干后,用掃描電鏡在150倍下檢測牙本質骨吸收陷窩,分別取三處,每處各照1張。用 Image J 計算骨吸收面積。
蛋白免疫印跡法
將 BMMs 接種于 6 孔板內過夜。對于短效蛋白(iκB -ɑ),加入氯化兩面針堿預孵育 1 小時,并加入 RANKL 按照0,10,20,30,60,120 分鐘時間點刺激;對于長效蛋白(NFATc1,D2),加入氯化兩面針堿與RANKL按照0,4,24,72,120,168 小時刺激。在檢測時間點后加入細胞裂解液,收集蛋白。
隨后配置 SDS-PAGE 凝膠,蛋白上樣,轉膜。用 5% 脫脂奶粉室溫封閉 1 小時后,TBST 洗膜,接下來分別孵育 iκB -ɑ 和 NFATc1,D2 抗體,最后 ECL 發光顯影。
構建去卵巢小鼠骨質疏松癥模型
C57BL/6J 小鼠實驗獲得西澳大學倫理委員會與廣西醫科大學倫理委員會批準(SCXK - (JUN) 2012-0004,Beijing, China)。 30 只 6-8 周齡的雌性小鼠隨機分為 5 組(即假手術組,對照組,藥物低劑量組,藥物高劑量組,雌激素組),每組各6只。除假手術組,每只老鼠腹腔注射 10% 水合氯醛后,雙側卵巢被摘除。術后恢復一周后,隔天給藥。根據體重和分組,進行腹腔注射。假手術組合對照組注射 1% DMSO,藥物低劑量組和高劑量組分別注射氯化兩面針堿(3mg/kg和 6mg/kg),雌激素組注射雌激素(50ng/kg)。給藥 6 周后,處死小鼠,并取其脛骨。脛骨用 4%多聚甲醛固定 24 小時后,用micro-CT掃描檢測。
分析
用 1×PBS 洗小鼠脛骨三次,裝入試管內掃描。應用 Bruker 1176 micro-CT 進行掃描,掃描條件為 50 KV-550 μA,0.5mm 鋁制濾波器,空間分辨率為 9 μM。用 NRecron 對數據進行重建 CT 圖;CTAn 分析軟件對樣品校準霍式值和密度。重用 CTAn 軟件(Bruker 公司)建后的樣本進行分析。以下是參數分析的方法:骨小梁和骨皮質的研究區是基于從生長板的底端來劃定。骨小梁的研究區是從生長板底端往下 0.5 毫米到 1.5 毫米區域。具有代表性的骨小梁研究區被通過畫曲線的方式從外圍的皮質骨中勾畫出來。骨小梁研究區通過恒定閾值被二元化,從而通過 CTAn 軟件進行量化分析。
骨組織形態學分析
用 4% 多聚甲醛固定脛骨 24 小時,1×PBS 沖洗 3 次。加入 14% EDTA 脫鈣 7 天。用石蠟包埋樣本,HE 染色或者 TRAcP 染色。通過 BIOQUANT OSTEO 軟件進行定量分析,獲取 OcS/BS 和 N.Oc/BS 等數據
統計學處理
所有實驗數據均重復 3 次。對于人外周血單個核細胞向破骨細胞分化過程中骨吸收功能實驗,結果按照均數±標準誤(x± SE)表示。通過 Excel 2003 軟件對各參數結果值進行處理分析,結果用均數±標準差(x± SEM)表示;假設檢驗設 P<0.05 具有統計學意義。
二、實驗結果
1)氯化兩面針堿對破骨細胞形成及前破骨細胞增殖、細胞凋亡的影響
選用來源于野生型小鼠的長骨骨髓中分離獲取的 BMMs 作為前破骨細胞,通過 RANKL(100ng/mL)和 M-CSF(10ng/mL)刺激誘導分化破骨細胞形成。RANKL(+) 為陽性對照組,RANKL(-)為空白組,其余加入不同濃度的氯化兩面針堿的進行培養(見圖 1-A)。通過 TRAcP 染色觀察,空白組無明顯破骨細胞形成,陽性對照組可見形態巨大、多核的破骨細胞。然而,氯化兩面針堿加入誘導體系中,隨著其濃度升高,對破骨細胞分化的抑制增強(見圖 1-B)。與陽性對照組相比,尤其在0.125 μM 到1 μM的氯化兩面針堿的作用下, 破骨細胞數顯著降低(見圖 1-C)。當藥物作用濃度超過 0.5 μM 時,幾乎完全沒有破骨細胞的形成。結果表明,氯化兩面針堿能夠抑制 RANKL 誘導的破骨細胞形成,且具有濃度依賴性。
為了證明氯化兩面針堿對破骨細胞形成的抑制作用不是通過對其凋亡而實現的,采用 Annexin V-FITC/PI 雙染色法,測定了不同濃度的氯化兩面針堿對前破骨細胞 RAW264.7 細胞凋亡的影響。結果顯示,不同濃度的氯化兩面針堿均沒有引起細胞的凋亡(見圖 1-D)。同時,采用 MTS 檢測氯化兩面針堿對前破骨細胞 BMMs 的增殖情況。結果顯示氯化兩面針堿作用濃度不超過 5 μM 時,對細胞的存活率沒有影響(見圖 1-E)。
2)氯化兩面針堿對RANKL誘導的破骨細胞分化相關基因表達的影響
采用 Real-Time PCR 檢測不同濃度氯化兩面針堿對 RANKL 誘導的BMMs 向破骨細胞分化 7 天后,相關基因 calcitonin receptor,cathepsin K,TRAcP,NFATc1,D2 表達。結果顯示,氯化兩面針堿能夠劑量依賴性的下調calcitonin receptor,cathepsin K,TRAcP,NFATc1,D2 mRNA 的表達(見圖 2)。結果在分子水平上證明了氯化兩面針堿對破骨細胞分化的抑制。
3)氯化兩面針堿對破骨細胞骨吸收功能的影響
破骨細胞是體內唯一具有骨吸收活性的細胞,頻繁過度的骨吸收活動是導致骨溶解疾病的根本原因。申請人將破骨樣細胞接種于骨板內,觀察氯化兩面針堿(0.5μM 和 1μM )對成熟破骨細胞骨吸收功能的影響。通過 TRAcP 染色觀察,與對照組相比,藥物沒有抑制已成熟破骨細胞的數量(見圖 3-A,B);然后觀察骨板,對照組可見大面積骨吸收陷窩;但是 1μM 氯化兩面針堿組可見少量骨吸收陷窩,且面積顯著減少(見圖 3-A,C)。結果表明氯化兩面針堿能夠抑制破骨細胞骨吸收功能。
4)氯化兩面針堿對誘導人外周血單個核細胞向破骨細胞分化相關基因的表達以及牙本質骨吸收功能的影響
選用人外周血單個核細胞作為破骨細胞前體,通過 hMCSF 與 hRANKL 共同誘導向破骨細胞分化。同時加入 1 μM 氯化兩面針堿進行孵育,考察其對人源破骨細胞的形成是否與鼠源破骨細胞形成的作用一致。 通過 TRAcP 染色發現,氯化兩面針堿顯著抑制成熟破骨細胞的標志物,cathepsin K,NFATc1,OSCAR, FcRγ, TREM2,DAP12 的基因表達(見圖 4-A)。
另外申請人還考察了氯化兩面針堿對人外周血單個核細胞向破骨細胞分化過程中骨吸收功能的影響。結果顯示,與對照組相比較,氯化兩面針堿(0.5 μM 和 1 μM)能夠顯著減少骨吸收陷窩面積。這與前文中發現氯化兩面針堿抑制鼠源破骨細胞活性結果一致(見圖 4-B,C)。
5)氯化兩面針堿對 RANKL 誘導的 NF-κB 活化的影響以及相關蛋白表達的調控
為了進一步探討氯化兩面針堿影響 RANKL 誘導破骨細胞分化的機制,申請人采用穩定轉染 3kB-Luc-SV40 熒光素酶報告基因的 Raw 264.7,檢測氯化兩面針堿對 RANKL 活化 NF-κB 的影響。結果顯示,100 ng/mL RANKL 能夠在 8 小時提高 NF-κB 的表達,提示 RANKL 能夠在體系中刺激 NF-κB 活化。并且發現當加入一系列濃度梯度的氯化兩面針堿時,能夠顯著降低 RANKL 誘導的 NF-κB 報告基因的表達,并且是隨著藥物濃度升高,抑制效果越明顯(見圖 5-A)。提示氯化兩面針堿有可能通過調控 NF-κB 的信號通路來影響破骨細胞的發生發展過程。
為了進一步闡明氯化兩面針堿對 NF-κB 信號通路作用,我們通過 Western-blot 檢測 NF-κB 活化的抑制因子 iκB -ɑ的蛋白表達情況。結果顯示,RANKL 誘導對照組能夠降低iκB -ɑ的蛋白表達,釋放 NF-κB 發揮作用。但是,氯化兩面針堿對 RANKL 刺激導致的 iκB -ɑ 蛋白降解沒有影響(見圖 5-B),提示氯化兩面針堿可能通過調控 iκB -ɑ 下游蛋白實現抑制 NF-κB 的信號通路。
6)氯化兩面針堿對 RANKL 誘導的 NFATc1 活化的影響以及相關蛋白表達的調控
NFATc1 目前被認為是破骨細胞關鍵的轉錄因子,能夠調節破骨細胞的分化,影響 calcitonin receptor,cathepsin K,TRAcP 等基因的表達。通過上述研究可知,氯化兩面針堿能夠下調上述基因 mRNA 的表達。因此,申請人通過熒光素酶報告基因與 Western-blot 考察了氯化兩面針堿對 RANKL 刺激 NFATc1 活化的影響。熒光素酶報告基因結果顯示,100 ng/mL RANKL 能夠在 24 小時顯著提高穩定轉染了 pNFATc1-TA-Luc 的 Raw264.7 中 NFATc1 的表達,提示 RANKL 能夠在這個體系中活化 NFATc1。當加入一系列濃度梯度的氯化兩面針堿時,顯著抑制了 RANKL 誘導的 NFATc1 報告基因表達的上升(見圖 6-A)。
Western-blot 結果發現,RANKL 誘導的 NFATc1,D2 蛋白水平顯著提高,加入氯化兩面針堿干預后,呈劑量依賴性降低 NFATc1 和 D2 的表達(見圖 6-B)。尤其在 1 μM 氯化兩面針堿干預下,不同時間點的 NFATc1 和 D2 的表達均被抑制(見圖 6-C)。提示氯化兩面針堿通過抑制轉錄因子 NFATc1 的表達,繼而調控其下游基因的表達,從而抑制破骨細胞的分化和功能。
7)氯化兩面針堿保護去卵巢骨質疏松小鼠的骨丟失
申請人已經通過體外 RANKL 誘導的破骨細胞體系發現氯化兩面針堿能夠抑制破骨細胞的形成和骨吸收功能。為了進一步證實氯化兩面針堿對破骨細胞的影響,申請人通過建立去卵巢小鼠骨質疏松癥模型,考察氯化兩面針堿對雌激素丟失引發過度骨吸收活動和破骨細胞的影響。通過 micro-CT 掃描檢測發現,建立去卵巢小鼠模型組 6 周后,出現明顯的骨丟失現象(見圖 7-A),骨小梁分離度明顯增加,骨體積分數、骨小梁厚度、以及骨小量數明顯降低(見圖 7-B)。從去卵巢小鼠骨頭組織形態學中發現,破骨細胞表面積/骨表面積與破骨細胞數目均降低(見圖 7-C)。而氯化兩面針堿高、低濃度劑量給藥組能夠有效的防止去卵巢小鼠導致的骨丟失(見圖 7-A),提升骨體積分數、骨小梁厚度、以及骨小量數,降低骨小梁分離度等骨靜態參數(見圖 7-B,C),其中高劑量給藥組(6mg/kg)的治療效果能夠類同于雌激素給藥組的功效(圖 7)。
三、討論
破骨細胞異常活躍的分化增殖和骨吸收功能容易引起骨丟失疾病,例如絕經性的骨質疏松癥,變形性骨炎,自身免疫性關節炎,牙周炎。這一過程通常是由 RANKL過度表達所致。RANKL 對破骨細胞前體啟動多種有關破骨細胞轉錄基因的胞內信號級聯放大,從而實現破骨細胞的分化與成熟。因此,靶標 RANKL 誘發的信號通路是治療骨丟失疾病的關注點。
通過體外實驗發現,氯化兩面針堿可以劑量依賴性抑制 RANKL 誘導的破骨細胞形成以及骨吸收功能作用。因此,詳細闡明氯化兩面針堿對破骨細胞的分子機制很有必要。破骨細胞前體向成熟的破骨細胞分化,需要多種細胞因子參與激活胞內信號通路。其中有兩個條件最為重要:一、MCSF,它是維持破骨細胞生長;二、RANKL,它是促進破骨細胞分化、成熟。目前,RANKL 引發的一系列變化與骨溶解疾病的發生發展緊密聯系。RANKL 聯合 RANK 作用于前破骨細胞,募集形成 TRAF6 與 c-src 復合蛋白體,并活化 IKK 激酶。IKK 通過降解 iκB-ɑ 來釋放 NF-κB 入核,與相應的 DNA 的啟動子結合,介導轉錄。我們發現,利用 NF-κB 熒光素酶報告基因,氯化兩面針堿劑量依賴性抑制 NF-κB 的表達。提示氯化兩面針堿可以通過 RANKL 誘導的 NF-κB 通路來抑制破骨細胞的形成及功能。但從蛋白免疫印跡結果顯示,氯化兩面針堿不能干預 RANKL 引起的 iκB-ɑ 降解,這說明氯化兩面針堿可能通過 iκB-ɑ 下游基因來抑制 NF-κB的活化。
RANKL 不僅能夠激活 NF-κB,還能夠通過 TRAF6 誘導活化轉錄因子 NFATc1 從而介導破骨細胞的成熟分化。通過 NFATc1 熒光素酶報告基因與蛋白免疫印跡發現,氯化兩面針堿也能夠抑制 NFATc1 的表達,從而影響破骨細胞的分化。并且下調了破骨細胞分化基因的表達;例如 cathepsin K,OSCAR,TREM2,FcRγ,DAP12 等。
為了進一步在體內證實氯化兩面針堿對破骨細胞的影響,通過建立去卵巢骨質疏松癥小鼠模型發現,氯化兩面針堿能夠保護去卵巢小鼠的骨丟失。與我們體外的研究結果一致。
綜上所述,氯化兩面針堿是未來防治破骨細胞引起的骨丟失疾病的一個理想藥物。
- 還沒有人留言評論。精彩留言會獲得點贊!