一種與致病力相關的灰霉病菌基因BcSEP5及其應用的制作方法
本發明屬微生物基因工程技術領域,具體涉及植物保護領域中控制真菌侵染結構形成與致病力的基因及其編碼蛋白質的應用。
背景技術:
灰霉病菌(Botrytis cinerea)通常又稱作灰葡萄孢,屬于子囊菌門(Ascomycota)真菌,是灰霉病的病原菌,可侵染400多種植物,包括幾乎所有的蔬菜及果樹作物。寄主從苗期到掛果期均可發病,而且,植株各部位均可被灰霉病菌侵染,葉部發病的典型癥狀表現為“V”形病斑,花部主要表現為腐爛和調萎,果實主要表現為腐爛和脫落。病害的發生和蔓延與環境的濕度、溫度存在密切的關系,在20℃-23℃,相對濕度90%以上時發生嚴重。因此,灰霉病屬低溫高濕型病害,在多雨季節或者保護地生產中極易發生,每年因該病導致的經濟損失高達100-1000億美元。由于寄主范圍廣泛,生產上危害嚴重,再加上相關分子研究技術成熟,灰霉菌已成為最重要的模式植物病原真菌之一,受到廣泛研究。
灰霉病菌是典型的死體營養型病原真菌,可生成多種致病因子參與致病,主要包括細胞壁降解酶類、角質酶、毒素、植物激素、抵抗寄主反應的酶類、小RNA及小分子物質等,這些因子相互協作使灰霉病菌能夠殺死寄主細胞并分解死亡的寄主組織作為營養。自然條件下,灰霉菌多以分生孢子作為侵染寄主的初侵染和再侵染來源。灰霉病菌常以菌絲體、分生孢子或菌核附著在植物病殘體上或在土壤中越冬和越夏,成為下一生長季的初侵染來源。當條件適宜時,菌核萌發產生菌絲體和分生孢子梗,并產生大量的分生孢子。成熟的分生孢子能夠借助風、雨水、灌溉水和農事操作等進行傳播。在低溫高濕條件下,分生孢子萌發形成芽管,芽管末端稍稍膨大發育成附著胞或者進一步形成侵染墊等侵染結構,主要從衰敗的花器、傷口以及壞死組織侵入。
附著胞、侵染墊等侵染結構的發育,對于灰霉病菌侵染寄主是至關重要的,如果侵染結構發育受到影響,灰霉病菌將難以侵入寄主,其危害程度隨之會受到嚴重削弱。灰霉病菌侵染結構的發育受到營養、界面等外源條件及發育信號的聯合調控,相關分子機制尚不清楚,對該領域進行深入研究不僅有助于揭示灰霉病菌等死體營養型病原真菌致病的分子機制,而且對于研發防治包括灰霉病菌在內的植物病原真菌的藥劑具有重要應用價值。
從分生孢子萌發到發育出功能完備的侵染結構,是一個精細的調控過程,鑒定該調控過程的重要組分,并驗證相關基因的致病功能,有可能從中發現可以作為殺真菌劑作用靶標的蛋白質,為開發防治灰霉病及其它類似病害的高效藥劑奠定理論和技術基礎。
Sep5是一種GTP結合蛋白質,廣泛存在于包括真菌在內的真核生物(植物除外),該蛋白質可通過聚合成異源寡聚體蛋白質復合體或進一步形成纖絲,參與調控細胞多種關鍵的生命過程。通過對灰霉病菌Sep5編碼基因的分析,評價該基因在灰霉病菌致病過程的作用,有利于鑒定潛在的防治靶標,用于篩選新型殺真菌藥劑。
技術實現要素:
本發明的目的旨在提供一種控制灰霉病菌侵染結構發育和致病性基因及其編碼的蛋白質。
本發明所提供的控制侵染結構發育和致病性基因來源于灰霉病菌,名稱為BcSEP5,其DNA序列如SEQ ID No:1所示。該DNA序列為BcSEP5基因開放閱讀框,由1571個核苷酸組成,其中包含7個外顯子,分別位于SEQ ID No:1的第1位至21位核苷酸之間,第163位至238位核苷酸之間,第296位至412位核苷酸之間,第474位至561位核苷酸之間,第612位至749位核苷酸之間,第803位至1270位核苷酸之間和第1322位至1571位核苷酸之間,組成的編碼區長度合計為1158個核苷酸。
本發明提供了來自灰霉病菌的控制侵染結構發育和致病性基因BcSEP5所編碼的蛋白質,其氨基酸序列如SEQ ID No:2所示,該序列由385個氨基酸組成。
來自灰霉病菌的控制侵染結構發育和致病性基因BcSEP5可應用于植物抗灰霉病基因工程領域。
來自灰霉病菌的控制侵染結構發育和致病性基因BcSEP5所編碼的蛋白質可作為靶標應用于抗灰霉病菌藥劑的設計與篩選。
本發明證明了BcSEP5基因的缺失或突變,導致灰霉病菌侵染結構形成率及致病力顯著降低,說明BcSEP5基因是灰霉病菌引起農作物灰霉病所必需的基因。因此,篩選能夠阻止該基因表達與其蛋白質的表達、修飾及定位的化合物,可以有效控制灰霉病的發生,從而有助于開發新型殺菌劑。即本發明所提供的BcSEP5基因的一個重要用途是:該基因的表達與其編碼的蛋白質產物,可以作為重要候選靶標位點,用于抗灰霉病菌藥劑的設計與篩選。
本發明所使用的灰霉病菌菌株B05.10,購買自美國真菌遺傳材料保藏中心(Fungal Genetics Stock Center,FGSC),其他人員如需要該菌株,可從該保藏中心購買獲得,相關信息如下:保藏中心地址:Fungal Genetics Stock Center,Department ofPlant Pathology,Kansas State University,4024 Throckmorton Plant Sciences Center,Manhattan,KS 66506USA;網址:http://www.fgsc.net/scripts/StrainSearchReturnPage.asp?OrgID=23812;菌株編號:FGSC 10317。
附圖說明
圖1為BcSEP5在灰霉病菌基因組中的基因結構圖譜
其中BcSEP5基因編號為BC1G_09606,預測該基因具有一個保守結構域:septin功能區——50s ribosome-binding GTPase。
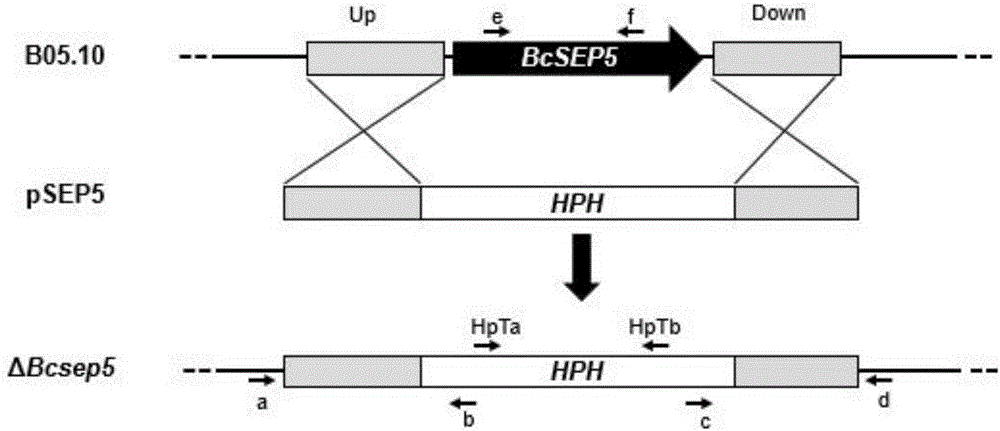
圖2為灰霉病菌BcSEP5基因的敲除策略(通過同源重組進行基因替換)示意圖
其中:B05.10為野生型菌株,pSEP5為敲除載體,ΔBcsep5為BcSEP5基因缺失突變體。a、b、c、d、e、f、HpTa、HpTb為驗證引物及其方向。
圖3為BcSEP5基因缺失突變體的PCR驗證電泳圖
其中:a、b、c、d、e、f、HpTa、HpTb為驗證引物,相應位置見圖2,a+b引物驗證基因上游是否發生同源交換,c+d引物驗證基因下游是否發生同源交換;e+f引物驗證目的基因是否被敲除。HpTa+HpTb引物驗證抗性篩選基因是否發生插入。ΔBcsep5-1和ΔBcsep5-2為兩株獨立獲得的BcSEP5基因缺失突變體。
圖4為敲除突變體ΔBcsep5與野生型菌株B05.10的培養特征對比照片
其中:所用基本培養基為PDA,B05.10為野生型,ΔBcsep5-1和ΔBcsep5-2為兩株獨立獲得的BcSEP5缺失突變體,RT-1和RT-2為隨機插入轉化子。
圖5為BcSEP5基因的缺失突變體與野生型菌株的產孢量統計分析
其中:B05.10為野生型,ΔBcsep5-1和ΔBcsep5-2為兩株獨立獲得的BcSEP5缺失突變體,RT-1為隨機插入轉化子,***代表具有顯著性差異。
圖6為BcSEP5基因的缺失突變體與野生型菌株的分生孢子形態差異比較
其中:B05.10為野生型,ΔBcsep5-1和ΔBcsep5-2為兩株獨立獲得的BcSEP5缺失突變體,RT-1為隨機插入轉化子,標尺為25μm。
圖7為BcSEP5基因的缺失突變體與野生型菌株的侵染墊發育對比分析
其中:B05.10為野生型,ΔBcsep5-1和ΔBcsep5-2為兩株獨立獲得的BcSEP5缺失突變體,RT-1為隨機插入轉化子,標尺為50μm。
圖8為BcSEP5基因的缺失突變體與野生型菌株致病力對比分析
其中:所選擇寄主為菜豆葉片,采用離體葉片接種方法,右側為接種菌絲塊,左側為接種孢子懸浮液,3天后進行評價。B05.10為野生型,ΔBcsep5-1和ΔBcsep5-2和ΔBcsep5-3為三株獨立獲得的BcSEP5缺失突變體,RT-1,RT-2為隨機插入轉化子。
具體實施方式
為了更好地描述本發明,下面通過具體的實施例予以進一步的說明,下述實施例中的方法,如無特別說明,均為常規方法。
實施例1 BcSEP5基因的相關性分析
灰霉病菌BcSEP5基因的開放閱讀框由1571個核苷酸組成,包含7個外顯子,編碼區cDNA全長為1158個核苷酸,編碼的蛋白質產物由385個氨基酸組成。對BcSEP5基因在灰霉病菌基因組網站(https://www.broadinstitute.org/fungal-genome-initiative/botrytis-cinerea-genome-project)中進行預測分析,BcSEP5基因編號為BC1G_09606,該基因具有一個保守結構域:septin功能區——50s ribosome-binding GTPase。
實施例2 BcSEP5基因的敲除與遺傳互補
1)敲除載體的構建
利用Primer 5.0設計引物,由蘇州金唯智生物技術有限公司進行引物合成,采用引物Sep5-UP-F(5'-GATCTTCACTAGTGGGAATTCCGTGCTCCAATGTTGTTTATCC-3')與Sep5-UP-R(5'-TTGGGTACCGAGCTCGAATTCAATCATGCGTCCCTCTTATCAA-3'),以灰霉病菌菌株B05.10的基因組DNA為模板擴增BcSEP5基因上游633bp片段;采用引物Sep5-DN-F(5'-TGGGGATCCTCTAGAGTCGACCTTTCGTTCGGATTTCTGGG-3')與Sep5-DN-R(5'-CTTGCATGCCTGCAGGTCGACTGGGAAGCGGGAGTGTATTT-3')擴增灰霉病菌BcSEP5基因下游627bp片段,反應體系(25μL)為:10mmol/L dNTP Mixture,0.5μL;10×PCRbuffer,2.5μL;上下游引物各1μL(10μmol/mL);模板DNA,1μL;Ex-Taq,0.2μL(5U);ddH2O,18.8μL;擴增程序為:94℃預變性3分鐘,然后(1)94℃,變性30秒;(2)60℃,退火30秒;(3)72℃,延伸60秒;(4)將1-3步循環30次;(5)72℃延伸10分鐘。將上述DNA擴增產物進行純化回收定量后,利用NEB生物公司的ClonExpress試劑盒,將上游片段和下游片段分別定向克隆至pXEH載體的EcoR I位點與Sal I位點,構建成敲除載體pSEP5(如圖2所示),并進行測序驗證。
2)灰霉病菌的轉化
a.根癌農桿菌的培養
挑取含有雙元載體pSEP5的根癌農桿菌菌株Agl-1單菌落,接種至含50μg/ml卡那霉素、10μg/ml利福平的MM液體培養基(磷酸氫二鉀0.205%,磷酸二氫鉀0.145%,氯化鈉0.015%,七水硫酸鎂0.05%,六水氯化鈣0.01%,七水硫酸亞鐵0.00025%,硫酸銨0.05%,葡萄糖0.2%)中,250rpm、28℃振蕩培養48h;4000rpm,離心5分鐘,棄上清,IM液體培養基(磷酸氫二鉀0.205%,磷酸二氫鉀0.145%,氯化鈉0.015%,七水硫酸鎂0.05%,六水氯化鈣0.01%,七水硫酸亞鐵0.00025%,硫酸銨0.05%,葡萄糖0.2%,200μMAS,MES 0.854%,甘油0.5%)重懸,4000rpm離心5分鐘,棄上清;IM培養基重懸,28℃,250rpm振蕩培養6h,進行預誘導。
b.灰霉病菌的產孢培養
選用B05.10菌株,取少量孢子涂布于PDA培養基(馬鈴薯20%煮爛過濾,葡萄糖2%,瓊脂1.5%),置28℃培養8h令孢子快速萌發,然后轉移至20℃培養3-5天,待菌體表面被灰色孢子覆蓋之后,用IM液體培養基刮取、收集孢子,顯微鏡觀察,利用血球計數器調節孢子濃度為1×106/mL。
c.根癌農桿菌與灰霉病菌分生孢子共培養及轉化子篩選
將在IM液體培養基中預先誘導6h的農桿菌菌液和灰霉病菌孢子液等體積混合,加入AS,使終濃度達到500μM,混勻,然后按250~350μL/皿,均勻涂抹至鋪有玻璃紙的IM培養基上,22℃黑暗培養48h;共培養完畢后,將玻璃紙轉移至含有100μg/mL潮霉素的PDA培養基上,相同條件下繼續培養。4~7天后挑取擴展的菌落到含有同樣抗生素的篩選培養基上。
3)缺失突變體的驗證
選用四對引物通過PCR擴增對轉化子進行篩選。擴增結果符合如下結果的確定為BcSEP5基因缺失突變體:潮霉素抗性基因內部引物HpTa(5'-TGCGCCCAAGCTGCATCAT-3')和HpTb(5'-TGAACTCACCGCGACGTCTGT-3')可以擴增到800bp片段(野生型菌株無擴增條帶);上游同源臂之外基因組上的引物a(5'-AGACCTACCTCTGGCAACTCG-3')與潮霉素抗性基因的引物b(5'-ACAGACGTCGCGGTGAGTTCA-3')配對可以擴增到預期(1.3kb)的重組片段;同時下游同源臂之外基因組上的引物d(5'-ATCTTATGCCGAACCTTCTGC-3')與潮霉素抗性基因的引物c(5'-ATGATGCAGCTTGGGCGCA-3')配對可以擴增到預期(0.9kb)的重組片段;而編碼區引物e(5'-ATCTTATGCCGAACCTTCTGC-3')與f(5'-GAGCTCGATATCTAATTCGCG-3')無擴增條帶(野生型菌株可擴增到0.4kb片段)。結果,從轉化子中篩選到2株獨立的BcSEP5基因缺失突變體:ΔBcsep5-1和ΔBcsep5-2(如圖3所示),用于后續功能分析。
實施例3 BcSEP5基因在灰霉病菌生長發育過程中的作用
采用平板培養法,評價BcSEP5突變體的生長發育等相關表型的變異情況。將待測菌株打取菌餅分別接種到含有100mg/L潮霉素的PDA培養基及不含有潮霉素的PDA培養基上,20℃黑暗培養3至5天后拍照觀察。結果發現,BcSEP5突變體的生長菌落直徑與野生型B05.10及隨機插入轉化子RT-1,RT-2相比嚴重滯后,而且BcSEP5突變體菌落(菌絲)形態發生明顯變化(見圖4),這說明BcSEP5基因在灰霉病菌正常的營養生長階段發揮重要功能,調控灰霉病菌的生長發育。
實施例4 BcSEP5基因在灰霉病菌產孢及分生孢子形態方面的作用
采用平板培養法,評價BcSEP5突變體的產孢能力的變異情況。將待測菌株接種到不含有潮霉素的PDA培養基上,20℃黑暗培養7至10天后,待菌絲長滿平板后開始產孢,用一定量的無菌水刮洗平板收集孢子懸浮液,并進行定量分析(見圖5),發現BcSEP5突變體的產孢量與野生型相比顯著下降;同時取10ul左右孢子懸浮液滴到載玻片上,制成臨時玻片,顯微觀察BcSEP5突變體的分生孢子形態(見圖6),發現BcSEP5突變體的分生孢子形態與野生型相比也發生了變異,孢子體形變小,由橢圓形變得更趨圓形;綜上所述,BcSEP5基因在灰霉病菌產孢及分生孢子形態建成方面也發揮重要作用。
實施例5 BcSEP5基因在灰霉病菌侵染結構——侵染墊發育過程中的作用
采用玻片培養法,觀察BcSEP5突變體發育成侵染墊的能力。將PDB孢子懸浮液(5×104mL-1)滴在載玻片上,室溫保濕培養36h。顯微觀察發現,野生型菌株B05.10及隨機插入轉化子RT的菌絲體可以形成大量多分枝墊狀結構——侵染墊,而突變體則幾乎喪失了發育成這種復雜侵染結構的能力;僅僅能形成一些零星的分枝較少的畸形侵染墊(見圖7)。該研究結果表明,BcSEP5基因在侵染墊發育過程中發揮舉足輕重的作用,該基因是灰霉病菌侵染結構發育所必需的。
實施例6 BcSEP5基因在灰霉病菌致病性方面的功能
采用離體葉片接種法,評價BcSEP5突變體的致病力變化情況。從溫室培養的寄主植株上采集具有一定葉齡的葉片,水平放至保濕容器中,將待測菌株進行產孢培養后,用PDB緩沖溶液收集分生孢子,并將孢子濃度調至2×106/mL,取5ul孢子懸浮液接種到葉片上,20保濕℃保濕黑暗培養,3天后評價待測菌株的致病力(見圖8左)。另外,使用打孔器打取待測菌株菌餅,正面朝下扣至葉片上,20℃保濕黑暗培養,3天后評價待測菌株的致病力(見圖8右)。實驗結果表明,不管是菌絲塊接種還是孢子懸浮液接種,BcSEP5突變體致病力均顯著下降,隨機插入轉化子致病力與野生型相似。綜上所述,BcSEP5是一個關鍵的致病基因,通過調控灰霉病菌侵染結構的發育影響其致病功能,如果該基因或其編碼蛋白質喪失活性,灰霉病菌侵染寄主引發病害的能力將顯著下降。
序 列 表
SEQ ID No:1的序列
(i)序列特征:(A)長度:1571 bp;(B)類型:核苷酸;(C)鏈性:單鏈
(ii)分子類型:DNA
(iii)序列描述:SEQ ID No:1
1 ATGTCGTCTC CATTCAATGC TGTATGTTTA TCCCTATCAA CAACTTTTTT
51 GGTCTATGTC CAAAGTCTGG ATAATCTGAC CGCAACTTCC CCAATAGTTC
101 ACATCTGGGG TTTGTTCTCA TCTTTCGAAG TTTGGTCAAG ACTAACTGAG
151 CAATTTCCCT AGATTAAGAT GCGAAAAAAG AAGAATGTCA AGAAGGGTAT
201 CCAGTTCTGT TTGATGGTCT GCGGTGCATC AGGCACAGGT ACGATAGCAC
251 AATGCACATA TGGATATCAT GGGCGGTATT GACTGGATCT CACAGGACGA
301 ACCACTTTCG TGAACACTTT ATGCGGCAAA CAGGTCCTCT CTGGCAAGGA
351 TGCCGACGAT GTGACAAATG CTCATGTCGA GGAAGGTGTC AAGATTAAGC
401 CAACCACAGT TGGTAAGCAA CAGTTGGATC TATAAAGTAT ATCTAAATTG
451 AACATTGACT GACACTCTTG CAGAGTTGGA ATTGGACGAG GAAGGAACCC
501 GTATTTCACT CACAATTGTC GACACACCTG GCTTTGGAGA CCAAATCGAC
551 AATGAGGCAA GGTAGGAGAG ACAAATTACA ACATGAGTGT TTCTAGCTGA
601 CAGATGTGAA GTTTTGGAGA GATTGTTGGC TACCTTGAGA GACAATACGA
651 CGATATTTTG GCAGAAGAAT CCCGTATCAA GCGTAATCCT CGTTTCCGCG
701 ACAATCGTAT CCATGCATTA CTTTATTTCA TCACCCCTAC CGGCCACGGG
751 TATGTCGCAT CTCAATCTTC ACCTCGCAAT TTCAAGCTGA CGTACTTCAT
801 AGCCTTCGCG AATTAGATAT CGAGCTCATG AAGCGTCTCT CCCCTAGAGT
851 TAATGTTATT CCAGTCATCG GAAAGGCTGA TTCCTTGACA CCCGCCGAAC
901 TTGCAGAATC AAAAAAGCTT GTCATGGAGG ATATCGAGCA CTATAGAATT
951 CCTGTCTACA ACTTCCCTTA TGATATCGAG GAGGACGACG AGGACACAGT
1001 TGAGGAGAAT GCTGAGCTTC GTGGTTTGAT GCCATTCGCC ATCGTGGGAT
1051 CAGAGGATGT TGTTGAGATT GGTGGACGTC AAGTTCGCGC AAGACAATAT
1101 CCATGGGGTG TTGTAGAGGT TGATAACCCA AGACATTCCG ACTTCCTTGC
1151 CATTAGAAGT GCCCTTCTAC ACAGTCATTT AGCAGATTTG AAGGAAATAA
1201 CCCACGATTT CCTTTACGAA AACTACCGTA CCGAGAAGCT TAGCAAGAGT
1251 GTTGACGGTG GTAGTGGAGG GTATGTTTTA TTCATTCTAA TTATGATTGA
1301 TAAATGCTAA TAGCTCTACA GTGATGCTTC TATGAACCCT GAGGATCTGG
1351 CCTCTCAATC TGTCCGCCTA AAGGAAGAGC AACTCCGCCG CGAGGAGGAG
1401 AAGCTTAGAG AGATTGAGAT CAAGGTGCAA CGAGAGATTG AGGCGAAGAG
1451 ACAAGAGTTA TTGGCTCGCG AGAGTCAATT AAAGGAGATC GAGGCTAGAA
1501 TGCAACGCGA GCAAACTGCT CATCTCGAGG CAACCAATGG AACACACACG
1551 TCAGCGGACG CTGAAGCTTA A
SEQ ID No:2的序列
(i)序列特征:(A)長度:385個氨基酸;(B)類型:氨基酸;(C)鏈性:單鏈
(ii)分子類型:多肽
(iii)序列描述:SEQ ID No:2
1 MSSPFNAIKM RKKKNVKKGI QFCLMVCGAS GTGRTTFVNT LCGKQVLSGK
51 DADDVTNAHV EEGVKIKPTT VELELDEEGT RISLTIVDTP GFGDQIDNEA
101 SFGEIVGYLE RQYDDILAEE SRIKRNPRFR DNRIHALLYF ITPTGHGLRE
151 LDIELMKRLS PRVNVIPVIG KADSLTPAEL AESKKLVMED IEHYRIPVYN
201 FPYDIEEDDE DTVEENAELR GLMPFAIVGS EDVVEIGGRQ VRARQYPWGV
251 VEVDNPRHSD FLAIRSALLH SHLADLKEIT HDFLYENYRT EKLSKSVDGG
301 SGGDASMNPE DLASQSVRLK EEQLRREEEK LREIEIKVQR EIEAKRQELL
351 ARESQLKEIE ARMQREQTAH LEATNGTHTS ADAEA
- 還沒有人留言評論。精彩留言會獲得點贊!