一種與致病力相關(guān)的灰霉病菌基因BcSEP6及其應(yīng)用的制作方法
本發(fā)明屬微生物基因工程技術(shù)領(lǐng)域,具體涉及植物保護(hù)領(lǐng)域中控制真菌侵染結(jié)構(gòu)形成與致病力的基因及其編碼蛋白質(zhì)的應(yīng)用。
背景技術(shù):
灰霉病菌(Botrytis cinerea)通常又稱(chēng)作灰葡萄孢,屬于子囊菌門(mén)(Ascomycota)真菌,是灰霉病的病原菌,可侵染400多種植物,包括幾乎所有的蔬菜及果樹(shù)作物。寄主從苗期到掛果期均可發(fā)病,而且,植株各部位均可被灰霉病菌侵染,葉部發(fā)病的典型癥狀表現(xiàn)為“V”形病斑,花部主要表現(xiàn)為腐爛和調(diào)萎,果實(shí)主要表現(xiàn)為腐爛和脫落。病害的發(fā)生和蔓延與環(huán)境的濕度、溫度存在密切的關(guān)系,在20℃-23℃,相對(duì)濕度90%以上時(shí)發(fā)生嚴(yán)重。因此,灰霉病屬低溫高濕型病害,在多雨季節(jié)或者保護(hù)地生產(chǎn)中極易發(fā)生,每年因該病導(dǎo)致的經(jīng)濟(jì)損失高達(dá)100-1000億美元。由于寄主范圍廣泛,生產(chǎn)上危害嚴(yán)重,再加上相關(guān)分子研究技術(shù)成熟,灰霉菌已成為最重要的模式植物病原真菌之一,受到廣泛研究。
灰霉病菌是典型的死體營(yíng)養(yǎng)型病原真菌,可生成多種致病因子參與致病,主要包括細(xì)胞壁降解酶類(lèi)、角質(zhì)酶、毒素、植物激素、抵抗寄主反應(yīng)的酶類(lèi)、小RNA及小分子物質(zhì)等,這些因子相互協(xié)作使灰霉病菌能夠殺死寄主細(xì)胞并分解死亡的寄主組織作為營(yíng)養(yǎng)。自然條件下,灰霉菌多以分生孢子作為侵染寄主的初侵染和再侵染來(lái)源。灰霉病菌常以菌絲體、分生孢子或菌核附著在植物病殘?bào)w上或在土壤中越冬和越夏,成為下一生長(zhǎng)季的初侵染來(lái)源。當(dāng)條件適宜時(shí),菌核萌發(fā)產(chǎn)生菌絲體和分生孢子梗,并產(chǎn)生大量的分生孢子。成熟的分生孢子能夠借助風(fēng)、雨水、灌溉水和農(nóng)事操作等進(jìn)行傳播。在低溫高濕條件下,分生孢子萌發(fā)形成芽管,芽管末端稍稍膨大發(fā)育成附著胞或者進(jìn)一步形成侵染墊等侵染結(jié)構(gòu),主要從衰敗的花器、傷口以及壞死組織侵入。
附著胞、侵染墊等侵染結(jié)構(gòu)的發(fā)育,對(duì)于灰霉病菌侵染寄主是至關(guān)重要的,如果侵染結(jié)構(gòu)發(fā)育受到影響,灰霉病菌將難以侵入寄主,其危害程度隨之會(huì)受到嚴(yán)重削弱。灰霉病菌侵染結(jié)構(gòu)的發(fā)育受到營(yíng)養(yǎng)、界面等外源條件及發(fā)育信號(hào)的聯(lián)合調(diào)控,相關(guān)分子機(jī)制尚不清楚,對(duì)該領(lǐng)域進(jìn)行深入研究不僅有助于揭示灰霉病菌等死體營(yíng)養(yǎng)型病原真菌致病的分子機(jī)制,而且對(duì)于研發(fā)防治包括灰霉病菌在內(nèi)的植物病原真菌的藥劑具有重要應(yīng)用價(jià)值。
從分生孢子萌發(fā)到發(fā)育出功能完備的侵染結(jié)構(gòu),是一個(gè)精細(xì)的調(diào)控過(guò)程,鑒定該調(diào)控過(guò)程的重要組分,并驗(yàn)證相關(guān)基因的致病功能,有可能從中發(fā)現(xiàn)可以作為殺真菌劑作用靶標(biāo)的蛋白質(zhì),為開(kāi)發(fā)防治灰霉病及其它類(lèi)似病害的高效藥劑奠定理論和技術(shù)基礎(chǔ)。
Sep6是一種GTP結(jié)合蛋白質(zhì),廣泛存在于包括真菌在內(nèi)的真核生物(植物除外),該蛋白質(zhì)可通過(guò)聚合成異源寡聚體蛋白質(zhì)復(fù)合體或進(jìn)一步形成纖絲,參與調(diào)控細(xì)胞多種關(guān)鍵的生命過(guò)程。通過(guò)對(duì)灰霉病菌Sep6編碼基因的分析,評(píng)價(jià)該基因在灰霉病菌致病過(guò)程的作用,有利于鑒定潛在的防治靶標(biāo),用于篩選新型殺真菌藥劑。
技術(shù)實(shí)現(xiàn)要素:
本發(fā)明的目的旨在提供一種控制灰霉病菌侵染結(jié)構(gòu)發(fā)育和致病性基因及其編碼的蛋白質(zhì)。
本發(fā)明所提供的控制侵染結(jié)構(gòu)發(fā)育和致病性基因來(lái)源于灰霉病菌,名稱(chēng)為BcSEP6,其DNA序列如SEQ ID No:1所示。該DNA序列為BcSEP6基因開(kāi)放閱讀框,由1345個(gè)核苷酸組成,其中包含4個(gè)外顯子,分別位于SEQ ID No:1的第1位至105位核苷酸之間,第169位至331位核苷酸之間,第397位至635位核苷酸之間和第764位至1345位核苷酸之間,組成的編碼區(qū)長(zhǎng)度合計(jì)為1089個(gè)核苷酸。
本發(fā)明提供了來(lái)自灰霉病菌的控制侵染結(jié)構(gòu)發(fā)育和致病性基因BcSEP6所編碼的蛋白質(zhì),其氨基酸序列如SEQ ID No:2所示,該序列由362個(gè)氨基酸組成。
來(lái)自灰霉病菌的控制侵染結(jié)構(gòu)發(fā)育和致病性基因BcSEP6可應(yīng)用于植物抗灰霉病基因工程領(lǐng)域。
來(lái)自灰霉病菌的控制侵染結(jié)構(gòu)發(fā)育和致病性基因BcSEP6所編碼的蛋白質(zhì)可作為靶標(biāo)應(yīng)用于抗灰霉病菌藥劑的設(shè)計(jì)與篩選。
本發(fā)明證明了BcSEP6基因的缺失或突變,導(dǎo)致灰霉病菌侵染結(jié)構(gòu)形成率及致病力顯著降低,說(shuō)明BcSEP6基因是灰霉病菌引起農(nóng)作物灰霉病所必需的基因。因此,篩選能夠阻止該基因表達(dá)與其蛋白質(zhì)的表達(dá)、修飾及定位的化合物,可以有效控制灰霉病的發(fā)生,從而有助于開(kāi)發(fā)新型殺菌劑。即本發(fā)明所提供的BcSEP6基因的一個(gè)重要用途是:該基因的表達(dá)與其編碼的蛋白質(zhì)產(chǎn)物,可以作為重要候選靶標(biāo)位點(diǎn),用于抗灰霉病菌藥劑的設(shè)計(jì)與篩選。
本發(fā)明所使用的灰霉病菌菌株B05.10,購(gòu)買(mǎi)自美國(guó)真菌遺傳材料保藏中心(Fungal Genetics Stock Center,F(xiàn)GSC),其他人員如需要該菌株,可從該保藏中心購(gòu)買(mǎi)獲得,相關(guān)信息如下:保藏中心地址:Fungal Genetics Stock Center,Department ofPlant Pathology,Kansas State University,4024Throckmorton Plant Sciences Center,Manhattan,KS 66506USA;網(wǎng)址:http://www.fgsc.net/scripts/StrainSearchReturnPage.asp?OrgID=23812;菌株編號(hào):FGSC 10317。
附圖說(shuō)明
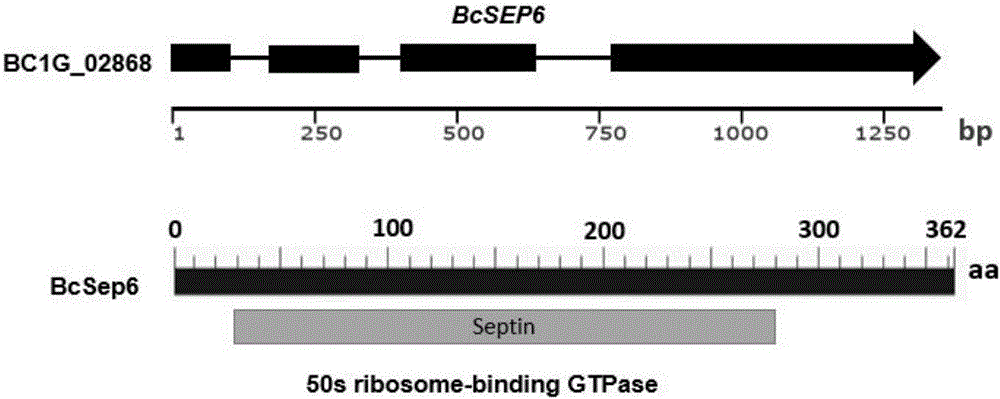
圖1為BcSEP6在灰霉病菌基因組中的基因結(jié)構(gòu)圖譜
其中BcSEP6基因編號(hào)為BC1G_02868,預(yù)測(cè)該基因具有一個(gè)GTPase結(jié)構(gòu)功能域
圖2為灰霉病菌BcSEP6基因的敲除策略(通過(guò)同源重組進(jìn)行基因替換)示意圖
其中:B05.10為野生型菌株,pSEP6為敲除載體,ΔBcsep6為BcSEP6基因缺失突變體。a、b、c、d、e、f、HpTa、HpTb為驗(yàn)證引物及其方向。
圖3為BcSEP6基因缺失突變體的PCR驗(yàn)證電泳圖
其中:a、b、c、d、e、f、HpTa、HpTb為驗(yàn)證引物,相應(yīng)位置見(jiàn)圖2,a+b引物驗(yàn)證基因上游是否發(fā)生同源交換,c+d引物驗(yàn)證基因下游是否發(fā)生同源交換;e+f引物驗(yàn)證目的基因是否被敲除。HpTa+HpTb引物驗(yàn)證抗性篩選基因是否發(fā)生插入。ΔBcsep6-1和ΔBcsep6-2為兩株獨(dú)立獲得的BcSEP6基因缺失突變體。RT為隨機(jī)插入轉(zhuǎn)化子
圖4為敲除突變體ΔBcsep6與野生型菌株B05.10的培養(yǎng)特征對(duì)比照片
其中:所用基本培養(yǎng)基為PDA,B05.10為野生型,ΔBcsep6-1和ΔBcsep6-2為兩株獨(dú)立獲得的BcSEP6缺失突變體,RT-1和RT-2為隨機(jī)插入轉(zhuǎn)化子。
圖5為BcSEP6基因的缺失突變體與野生型菌株的產(chǎn)孢量統(tǒng)計(jì)分析
其中:B05.10為野生型,ΔBcsep6-1和ΔBcsep6-2為兩株獨(dú)立獲得的BcSEP6缺失突變體,RT-1為隨機(jī)插入轉(zhuǎn)化子,***代表具有顯著性差異。
圖6為BcSEP6基因的缺失突變體與野生型菌株的分生孢子形態(tài)差異比較
其中:B05.10為野生型,ΔBcsep6-1和ΔBcsep6-2為兩株獨(dú)立獲得的BcSEP6缺失突變體,RT-1為隨機(jī)插入轉(zhuǎn)化子,標(biāo)尺為25μm。
圖7為BcSEP6基因的缺失突變體與野生型菌株的侵染墊發(fā)育對(duì)比分析
其中:B05.10為野生型,ΔBcsep6-1和ΔBcsep6-2為兩株獨(dú)立獲得的BcSEP6缺失突變體,RT-1為隨機(jī)插入轉(zhuǎn)化子,標(biāo)尺為50μm。
圖8為BcSEP6基因的缺失突變體與野生型菌株致病力對(duì)比分析
其中:所選擇寄主為菜豆葉片,采用離體葉片接種方法,左側(cè)為接種菌絲塊,右側(cè)為接種孢子懸浮液,3天后進(jìn)行評(píng)價(jià)。B05.10為野生型,ΔBcsep6-1和ΔBcsep6-2和ΔBcsep6-3為三株獨(dú)立獲得的BcSEP6缺失突變體,RT-1,RT-2為隨機(jī)插入轉(zhuǎn)化子;dpi表示接種后天數(shù)。
具體實(shí)施方式
為了更好地描述本發(fā)明,下面通過(guò)具體的實(shí)施例予以進(jìn)一步的說(shuō)明,下述實(shí)施例中的方法,如無(wú)特別說(shuō)明,均為常規(guī)方法。
實(shí)施例1 BcSEP6基因的相關(guān)性分析
灰霉病菌BcSEP6基因的開(kāi)放閱讀框由1345個(gè)核苷酸組成,包含4個(gè)外顯子,編碼區(qū)cDNA全長(zhǎng)為1089個(gè)核苷酸,編碼的蛋白質(zhì)產(chǎn)物由362個(gè)氨基酸組成。對(duì)BcSEP6基因在灰霉病菌基因組網(wǎng)站(https://www.broadinstitute.org/fungal-genome-initiative/botrytis-cinerea-genome-project)中進(jìn)行預(yù)測(cè)分析,BcSEP6基因編號(hào)為BC1G_02868,該基因具有一個(gè)保守GTPase結(jié)構(gòu)功能域。
實(shí)施例2 BcSEP6基因的敲除與遺傳互補(bǔ)
1)敲除載體的構(gòu)建
利用Primer 5.0設(shè)計(jì)引物,由蘇州金唯智生物技術(shù)有限公司進(jìn)行引物合成,采用引物Sep6-UP-F(5'-GATCTTCACTAGTGGGAATTCTCGATGTGCGAATGGAAGTG-3')與Sep6-UP-R(5'-TTGGGTACCGAGCTCGAATTCGCTGTCGGTGGGTTTGTGGT-3'),以灰霉病菌菌株B05.10的基因組DNA為模板擴(kuò)增BcSEP6基因上游600bp片段;采用引物Sep6-DN-F(5'-TGGGGATCCTCTAGAGTCGACCCACGATGTAATTCATGCTCCT-3')與Sep6-DN-R(5'-CTTGCATGCCTGCAGGTCGACATCGGTGAACTACTACGAAACTCC-3')擴(kuò)增灰霉病菌BcSEP6基因下游809bp片段,反應(yīng)體系(25μL)為:10mmol/L dNTP Mixture,0.5μL;10×PCR buffer,2.5μL;上下游引物各1μL(10μmol/mL);模板DNA,1μL;Ex-Taq,0.2μL(5U);ddH2O,18.8μL;擴(kuò)增程序?yàn)椋?4℃預(yù)變性3分鐘,然后(1)94℃,變性30秒;(2)58℃,退火30秒;(3)72℃,延伸60秒;(4)將1-3步循環(huán)30次;(5)72℃延伸10分鐘。將上述DNA擴(kuò)增產(chǎn)物進(jìn)行純化回收定量后,利用NEB生物公司的ClonExpress試劑盒,將上游片段和下游片段分別定向克隆至pXEH載體的EcoR I位點(diǎn)與Sal I位點(diǎn),構(gòu)建成敲除載體pSEP6(如圖2所示),并進(jìn)行測(cè)序驗(yàn)證。
2)灰霉病菌的轉(zhuǎn)化
a.根癌農(nóng)桿菌的培養(yǎng)
挑取含有雙元載體pSEP6的根癌農(nóng)桿菌菌株Agl-1單菌落,接種至含50μg/ml卡那霉素、10μg/ml利福平的MM液體培養(yǎng)基(磷酸氫二鉀0.205%,磷酸二氫鉀0.145%,氯化鈉0.015%,七水硫酸鎂0.05%,六水氯化鈣0.01%,七水硫酸亞鐵0.00025%,硫酸銨0.05%,葡萄糖0.2%)中,250rpm、28℃振蕩培養(yǎng)48h;4000rpm,離心5分鐘,棄上清,IM液體培養(yǎng)基(磷酸氫二鉀0.205%,磷酸二氫鉀0.145%,氯化鈉0.015%,七水硫酸鎂0.05%,六水氯化鈣0.01%,七水硫酸亞鐵0.00025%,硫酸銨0.05%,葡萄糖0.2%,200μMAS,MES 0.854%,甘油0.5%)重懸,4000rpm離心5分鐘,棄上清;IM培養(yǎng)基重懸,28℃,250rpm振蕩培養(yǎng)6h,進(jìn)行預(yù)誘導(dǎo)。
b.灰霉病菌的產(chǎn)孢培養(yǎng)
選用B05.10菌株,取少量孢子涂布于PDA培養(yǎng)基(馬鈴薯20%煮爛過(guò)濾,葡萄糖2%,瓊脂1.5%),置28℃培養(yǎng)8h令孢子快速萌發(fā),然后轉(zhuǎn)移至20℃培養(yǎng)3-5天,待菌體表面被灰色孢子覆蓋之后,用IM液體培養(yǎng)基刮取、收集孢子,顯微鏡觀察,利用血球計(jì)數(shù)器調(diào)節(jié)孢子濃度為1×106/mL。
c.根癌農(nóng)桿菌與灰霉病菌分生孢子共培養(yǎng)及轉(zhuǎn)化子篩選
將在IM液體培養(yǎng)基中預(yù)先誘導(dǎo)6h的農(nóng)桿菌菌液和灰霉病菌孢子液等體積混合,加入AS,使終濃度達(dá)到500μM,混勻,然后按250~350μL/皿,均勻涂抹至鋪有玻璃紙的IM培養(yǎng)基上,22℃黑暗培養(yǎng)48h;共培養(yǎng)完畢后,將玻璃紙轉(zhuǎn)移至含有100μg/mL潮霉素的PDA培養(yǎng)基上,相同條件下繼續(xù)培養(yǎng)。4~7天后挑取擴(kuò)展的菌落到含有同樣抗生素的篩選培養(yǎng)基上。
3)缺失突變體的驗(yàn)證
選用四對(duì)引物通過(guò)PCR擴(kuò)增對(duì)轉(zhuǎn)化子進(jìn)行篩選。擴(kuò)增結(jié)果符合如下結(jié)果的確定為BcSEP6基因缺失突變體:潮霉素抗性基因內(nèi)部引物HpTa(5'-TGCGCCCAAGCTGCATCAT-3')和HpTb(5'-TGAACTCACCGCGACGTCTGT-3')可以擴(kuò)增到800bp片段(野生型菌株無(wú)擴(kuò)增條帶);上游同源臂之外基因組上的引物a(5'-TCATTCGTCCAACTCCCACAG-3')與潮霉素抗性基因的引物b(5'-ACAGACGTCGCGGTGAGTTCA-3')配對(duì)可以擴(kuò)增到預(yù)期(1.2kb)的重組片段;同時(shí)下游同源臂之外基因組上的引物d(5'-CAACAAACTCAGACTCCGCTG-3')與潮霉素抗性基因的引物c(5'-ATGATGCAGCTTGGGCGCA-3')配對(duì)可以擴(kuò)增到預(yù)期(1.2kb)的重組片段;而編碼區(qū)引物e(5'-GAATCTGGACTTGGAAAGAC-3')與f(5'-CTTCCTAGGTCTAGCTTCAC-3')無(wú)擴(kuò)增條帶(野生型菌株可擴(kuò)增到0.94kb片段)。結(jié)果,從轉(zhuǎn)化子中篩選到2株獨(dú)立的BcSEP6基因缺失突變體:ΔBcsep6-1和ΔBcsep6-2(如圖3所示),用于后續(xù)功能分析。
實(shí)施例3 BcSEP6基因在灰霉病菌生長(zhǎng)發(fā)育過(guò)程中的作用
采用平板培養(yǎng)法,評(píng)價(jià)BcSEP6突變體的生長(zhǎng)發(fā)育等相關(guān)表型的變異情況。將待測(cè)菌株打取菌餅分別接種到含有100mg/L潮霉素的PDA培養(yǎng)基及不含有潮霉素的PDA培養(yǎng)基上,20℃黑暗培養(yǎng)3至5天后拍照觀察。結(jié)果發(fā)現(xiàn),BcSEP6突變體的生長(zhǎng)菌落直徑與野生型B05.10及隨機(jī)插入轉(zhuǎn)化子RT-1,RT-2相比嚴(yán)重滯后,而且BcSEP6突變體菌落(菌絲)形態(tài)發(fā)生明顯變化(見(jiàn)圖4),這說(shuō)明BcSEP6基因在灰霉病菌正常的營(yíng)養(yǎng)生長(zhǎng)階段發(fā)揮重要功能,調(diào)控灰霉病菌的生長(zhǎng)發(fā)育。
實(shí)施例4 BcSEP6基因在灰霉病菌產(chǎn)孢及分生孢子形態(tài)方面的作用
采用平板培養(yǎng)法,評(píng)價(jià)BcSEP6突變體的產(chǎn)孢能力的變異情況。將待測(cè)菌株接種到不含有潮霉素的PDA培養(yǎng)基上,20℃黑暗培養(yǎng)7至10天后,待菌絲長(zhǎng)滿(mǎn)平板后開(kāi)始產(chǎn)孢,用一定量的無(wú)菌水刮洗平板收集孢子懸浮液,并進(jìn)行定量分析(見(jiàn)圖5),發(fā)現(xiàn)BcSEP6突變體的產(chǎn)孢量與野生型相比顯著下降;同時(shí)取10ul左右孢子懸浮液滴到載玻片上,制成臨時(shí)玻片,顯微觀察BcSEP6突變體的分生孢子形態(tài)(見(jiàn)圖6),發(fā)現(xiàn)BcSEP6突變體的分生孢子形態(tài)與野生型相比也發(fā)生了變異,孢子體形變小,由橢圓形變得更趨圓形;綜上所述,BcSEP6基因在灰霉病菌產(chǎn)孢及分生孢子形態(tài)建成方面也發(fā)揮重要作用。
實(shí)施例5 BcSEP6基因在灰霉病菌侵染結(jié)構(gòu)——侵染墊發(fā)育過(guò)程中的作用
采用玻片培養(yǎng)法,觀察BcSEP6突變體發(fā)育成侵染墊的能力。將PDB孢子懸浮液(5×104mL-1)滴在載玻片上,室溫保濕培養(yǎng)36h。顯微觀察發(fā)現(xiàn),野生型菌株B05.10及隨機(jī)插入轉(zhuǎn)化子RT的菌絲體可以形成大量多分枝墊狀結(jié)構(gòu)——侵染墊,而突變體則幾乎喪失了發(fā)育成這種復(fù)雜侵染結(jié)構(gòu)的能力;僅僅能形成一些零星的分枝較少的畸形侵染墊(見(jiàn)圖7)。該研究結(jié)果表明,BcSEP6基因在侵染墊發(fā)育過(guò)程中發(fā)揮舉足輕重的作用,該基因是灰霉病菌侵染結(jié)構(gòu)發(fā)育所必需的。
實(shí)施例6 BcSEP6基因在灰霉病菌致病性方面的作用
采用離體葉片接種法,評(píng)價(jià)BcSEP6突變體的致病力變化情況。從溫室培養(yǎng)的寄主植株上采集具有一定葉齡的葉片,水平放至保濕容器中,使用打孔器打取待測(cè)菌株菌餅,正面朝下扣至葉片上,20℃保濕黑暗培養(yǎng),3天后評(píng)價(jià)待測(cè)菌株的致病力(見(jiàn)圖8左)。另外,將待測(cè)菌株進(jìn)行產(chǎn)孢培養(yǎng)后,用PDB緩沖溶液收集分生孢子,并將孢子濃度調(diào)至2×106/mL,取5ul孢子懸浮液接種到葉片上,20℃保濕黑暗培養(yǎng),3天后評(píng)價(jià)待測(cè)菌株的致病力(見(jiàn)圖8右)。實(shí)驗(yàn)結(jié)果表明,不管是菌絲塊接種還是孢子懸浮液接種,BcSEP6突變體致病力均顯著下降,隨機(jī)插入轉(zhuǎn)化子致病力與野生型相似,綜上研究表明,BcSEP6是一個(gè)關(guān)鍵的致病基因,在灰霉病菌侵染寄主過(guò)程中發(fā)揮重要作用。BcSEP6通過(guò)調(diào)控灰霉病菌侵染結(jié)構(gòu)的發(fā)育,進(jìn)而影響其致病功能,如果該基因或其編碼蛋白質(zhì)喪失活性,灰霉病菌侵染寄主引發(fā)病害的能力將嚴(yán)重下降。
序 列 表
SEQ ID No:1的序列
(i)序列特征:(A)長(zhǎng)度:1345 bp;(B)類(lèi)型:核苷酸;(C)鏈性:?jiǎn)捂?/p>
(ii)分子類(lèi)型:DNA
(iii)序列描述:SEQ ID No:1
1 ATGGCACCAC CCACCGCCGA AAGCGCTTCC CCCATCGGAA TTGCGAATTT
51 GCCCAACCAA AGACACAAGA TTGTCGCAAA GAGAGGTGCT GCCTTCACTA
101 TTATGGTATG TGGTTATCTA TGCTGCTTTC AAATCTCTCA GCAACTTTAT
151 ACTAATCACC TCTTGTAGGT CGCTGGCGAA TCTGGACTTG GAAAGACAAC
201 CTTCATTAAT ACCCTCTTCT CTACCACAAT CAAAAACTAT GCCGATCACA
251 AACGCAGACA CGCAAAGCAA GTGGATAAAA CTGTTGAGAT TGAAATCACA
301 AAGGCTGAAT TAGAAGAGAA ATTCTTCAAG GGTATGGAAT CCAACACATT
351 CAGTTCACTT GAGATAATTA ATTGGCCATT GTTGACACTG TCTTAGTTCG
401 TTTGACTGTT ATCGATACAC CTGGTTTTGG AGATTATGTC AACAATCGCG
451 ACTCCTGGAT GCCAATCATC GAGTTTTTGG ATGATCAACA TGAGTCCTAT
501 ATGTTGCAAG AGCAACAGCC TCGCCGTGTT GATAAAATCG ATTTACGTGT
551 ACACGCTTGC TTGTACTTCA TCCGACCTAC TGGCCACACA CTGAAACCCC
601 TCGATATTGA GGTTATGAAG AGACTCAGTT CAAGGGTAAA CTTGATTCCA
651 GTAGTTGCTA AGGCTGATAC CTTAAGCCCG GCTGATTTGG CCCGTTATAA
701 GCAAAAGGTA TAGTGTATTT GAGTAAACCA TTATCGTTGT CGAAGTTAAC
751 GTATTTTCTA CAGATCCAAG CTGTCATTGA AGCACAAGGT ATCAAGATCT
801 ACACACCACC TATCGAGGAA GATGACGAGG CTGCTGCTCA ACATGCCCGA
851 AGTTTGATGG CTGCTATGCC ATTTGCAGTC ATTGGCTCAG AGAAGGATGT
901 CAAGACTAGT GATGGACGTA TCGTTAAGGG TCGTCAATAC GCATGGGGTG
951 TTGCTGAAGT CGAGAACGAA GACCACTGCG ATTTCAAGAA GTTGAGATCC
1001 ATTCTTATCC GAACACACAT GCTCGATCTC ATTCACACTA CCGAAGAAGC
1051 ACACTACGAG GCTTACCGCG CCCAACAAAT GGAGACTCGC AAATTCGGTG
1101 AAGCTAGACC TAGGAAGCTT GATAACCCTA AATTCAAGGA AGAGGAAGAG
1151 AGCTTAAGAA AGCGTTTCAC CGAGCAGGTT AAGATTGAGG AGCACCGTTT
1201 CAGACAGTGG GAGCAGAAGC TCATCAGTGA GCGTGATCGT TTGAACAAGG
1251 ATTTGGAGAG TACACATGCT GCCATCAAGT CCTTAGAACA AGAACTTGAG
1301 CAAATGCAAG GCGGTGCCGC AACTCGCAGT CACGGCCGTC GTTAA
SEQ ID No:2的序列
(i)序列特征:(A)長(zhǎng)度:362個(gè)氨基酸;(B)類(lèi)型:氨基酸;(C)鏈性:?jiǎn)捂湣?/p>
(ii)分子類(lèi)型:多肽
(iii)序列描述:SEQ ID No:2
1 MAPPTAESAS PIGIANLPNQ RHKIVAKRGA AFTIMVAGES GLGKTTFINT
51 LFSTTIKNYA DHKRRHAKQV DKTVEIEITK AELEEKFFKV RLTVIDTPGF
101 GDYVNNRDSW MPIIEFLDDQ HESYMLQEQQ PRRVDKIDLR VHACLYFIRP
151 TGHTLKPLDI EVMKRLSSRI QAVIEAQGIK IYTPPIEEDD EAAAQHARSL
201 MAAMPFAVIG SEKDVKTSDG RIVKGRQYAW GVAEVENEDH CDFKKLRSIL
251 IRTHMLDLIH TTEEAHYEAY RAQQMETRKF GEARPRKLDN PKFKEEEESL
301 RKRFTEQVKI EEHRFRQWEQ KLISERDRLN KDLESTHAAI KSLEQELEQM
351 QGGAATRSHG RR
- 還沒(méi)有人留言評(píng)論。精彩留言會(huì)獲得點(diǎn)贊!
- 一株體外高效拮抗稻瘟病菌的水稻內(nèi)生放線菌OsiLf?2的制作方法與工藝
- 一種用于篩選拮抗菌的刻度尺的制作方法與工藝
- 一種灰霉病菌拮抗菌株及其篩選方法和應(yīng)用與流程
- 一種高生防力葡萄灰霉菌拮抗菌的快速誘抗篩選方法與流程
- 一種與致病力相關(guān)的灰霉病菌基因BcSEP5及其應(yīng)用的制作方法與工藝
- 一種與致病力相關(guān)的灰霉病菌基因BcSEP3及其應(yīng)用的制作方法與工藝
- 一種與致病力相關(guān)的灰霉病菌基因BcPCK1及其應(yīng)用的制作方法與工藝
- 一種防治番茄灰霉病的生物殺菌劑及其制備方法與流程
- 一種防治大棚番茄灰霉病的煙劑及其制備方法與流程
- 一種灰霉病菌對(duì)多種藥劑抗藥性的快速檢測(cè)方法與流程
- 馬鈴薯瘡痂病菌定量分子檢測(cè)方法
- 基于大數(shù)據(jù)群體行為的疾病相似度計(jì)算方法及裝置的制造方法
- 一種基于二代測(cè)序檢測(cè)Rett綜合癥致病基因SNP位點(diǎn)的試劑盒及其檢測(cè)方法
- 一種治療肝內(nèi)膽汁淤積性黃疸疾病的中藥組合物及其應(yīng)用
- 檢測(cè)耳聾基因的核酸序列及其應(yīng)用
- 骨硬化癥新的突變致病基因plekhm1、及其編碼蛋白和應(yīng)用
- 一種富集4000人類(lèi)致病靶基因的方法及試劑盒的制作方法
- 一種黃曲霉致病基因AfsumO及應(yīng)用
- 一種基因傳遞系統(tǒng)及其制備方法
- 白化病ii型致病基因突變檢測(cè)試劑盒的制作方法