免疫應(yīng)答的引發(fā)的制作方法
本申請(qǐng)是國際申請(qǐng)日為2009年11月20日、國際申請(qǐng)?zhí)枮閜ct/dk2009/050310、進(jìn)入國家階段的申請(qǐng)?zhí)枮?00980154924.4、發(fā)明名稱為“免疫應(yīng)答的引發(fā)”的pct申請(qǐng)的分案申請(qǐng)。
本申請(qǐng)中引用的所有專利和非專利參考文獻(xiàn)在此通過引用整體并入。
本發(fā)明涉及使用恒定鏈連接的抗原的引發(fā)免疫應(yīng)答的技術(shù)和方法(當(dāng)這些用于引發(fā)使用任何合適疫苗的后續(xù)加強(qiáng)免疫接種時(shí))。
背景技術(shù):
疫苗接種是給受試者施用抗原材料(疫苗),以便產(chǎn)生對(duì)疾病或狀況的免疫。當(dāng)用于刺激免疫應(yīng)答時(shí),抗原被稱為免疫原,并且該過程被稱為免疫接種。疫苗接種涉及一種或多種免疫原的施用,這可以以幾種形式施用。
疫苗接種要求實(shí)體免疫應(yīng)答的建立。由感染或疫苗接種激活的免疫應(yīng)答依賴于幾種細(xì)胞類型例如t、b和抗原呈遞細(xì)胞,以及幾種不同分子主要是抗原、mhc分子、t和b細(xì)胞受體及許多其他的相互作用。
常規(guī)疫苗或第一代疫苗基于被殺死或減毒的致病菌株。這些通常具有減少的傳染力,并且它們通常是無足夠的免疫原性的,導(dǎo)致來自疫苗接種的不充分保護(hù)。
基于來自致病生物的個(gè)別抗原蛋白質(zhì)的第二代疫苗或亞單位疫苗的開發(fā)已揭示純肽或碳水化合物趨于是弱免疫原。
dna疫苗或第三代疫苗具有誘導(dǎo)較廣泛范圍的免疫應(yīng)答類型的能力,但維持在人中具有低免疫原性的潛在缺點(diǎn)。
對(duì)于所有類型的疫苗,疫苗接種程序面對(duì)關(guān)于增加疫苗效力的未滿足需要,以克服上述局限性且提供在疫苗接種過程中刺激免疫系統(tǒng)的更經(jīng)濟(jì)方法。
本發(fā)明目標(biāo)在于通過提供用于增加疫苗效力的解決方案來解決與疫苗的低免疫原性相關(guān)的問題。
本發(fā)明目標(biāo)在于通過提供用于引發(fā)免疫應(yīng)答的解決方案來解決與疫苗的低免疫原性相關(guān)的問題。
本發(fā)明因此涉及用核酸構(gòu)建體引發(fā)免疫系統(tǒng),所述核酸構(gòu)建體包括mhcii類結(jié)合的恒定鏈/cd74(本文被稱為恒定鏈或ii)或其變體,且編碼至少一種抗原蛋白質(zhì)或所述抗原蛋白質(zhì)的片段,隨后為后續(xù)加強(qiáng)疫苗接種,以增加所述疫苗的效力。
根據(jù)本發(fā)明的疫苗可以涉及致病性抗原或癌抗原。
本文呈現(xiàn)的數(shù)據(jù)顯示使用包括恒定鏈或其變體的核酸構(gòu)建體開發(fā)引發(fā)-加強(qiáng)方案并不簡單。
令人驚訝的是,本發(fā)明公開了可以改變來自恒定鏈的ii-key(包括lrmk氨基酸殘基)和/或ii-clip結(jié)構(gòu)域的部分,而不減少所述免疫引發(fā)的效應(yīng)。
不充分地解決這個(gè)問題的現(xiàn)有技術(shù)參考文獻(xiàn)在下文討論:
wo2007/062656(holst等人)涉及開發(fā)改良dna疫苗,以這樣的方式刺激免疫應(yīng)答,所述方式增加應(yīng)答的動(dòng)力學(xué),同時(shí)伴隨擴(kuò)大且改善應(yīng)答。holst等人發(fā)現(xiàn)抗原與恒定鏈的融合通過cd4+t細(xì)胞獨(dú)立機(jī)制顯著增強(qiáng)隨后的抗病毒cd4+和cd8+t細(xì)胞應(yīng)答。
holmes等人(jclinoncol.2008,jul10;26(20):3426-33)描述了ii-key雜合肽疫苗的首次人i期試驗(yàn),其中ii-key包括lrmk4氨基酸序列,恒定鏈蛋白質(zhì)的中心部分。
kallinteris等人的發(fā)現(xiàn)(expertopin.biol.ther.2006,6(12):1311,1321)也涉及利用包括lrmk氨基酸的ii-key部分用于增強(qiáng)疫苗效力。
us2008/0095798(humphreys等人)公開了用于增加疫苗針對(duì)病原體的效力的方法,通過首先用包括所述ii-key肽的lrmk殘基的ii-key雜合肽構(gòu)建體引發(fā)受試者的免疫系統(tǒng),并且隨后施用針對(duì)病原體的疫苗以加強(qiáng)在引發(fā)步驟中產(chǎn)生的免疫應(yīng)答。
技術(shù)實(shí)現(xiàn)要素:
本發(fā)明顯示用核酸構(gòu)建體引發(fā)免疫系統(tǒng),所述核酸構(gòu)建體包括至少一種mhcii類結(jié)合的恒定鏈/cd74(本文被稱為恒定鏈或ii)或其變體,且編碼至少一種抗原蛋白質(zhì)或所述抗原蛋白質(zhì)的片段,隨后為后續(xù)加強(qiáng)疫苗接種增加所述疫苗的效力。
根據(jù)本發(fā)明的疫苗可以涉及致病性抗原或癌抗原。
令人驚訝的是,本發(fā)明公開了ii-key結(jié)構(gòu)域(包括lrmk氨基酸殘基)和/或ii-clip結(jié)構(gòu)域的部分可以從恒定鏈中部分置換或省略,而不降低所述引發(fā)的作用。
因此,本發(fā)明已解決了通過采用二步引發(fā)-加強(qiáng)方案的疫苗接種而產(chǎn)生的充分刺激免疫應(yīng)答的問題,由此首先用包括恒定鏈或其變體的核酸構(gòu)建體引發(fā)免疫系統(tǒng),隨后為以這樣的方式使用任何類型的合適疫苗的后續(xù)加強(qiáng)免疫接種,所述方式增加應(yīng)答的動(dòng)力學(xué),同時(shí)伴隨擴(kuò)大且改善應(yīng)答。特別地,在此可提供用于直接、特異和快速刺激免疫系統(tǒng)的新型系統(tǒng),以便改善所有動(dòng)物例如人的疫苗接種方案。
這個(gè)問題通過在權(quán)利要求中表征的本發(fā)明的實(shí)施方案得到解決。通過本發(fā)明,發(fā)現(xiàn)用基于ii鏈的核酸構(gòu)建體的引發(fā)顯著增加在用疫苗后續(xù)加強(qiáng)后的抗原特異性cd8+t細(xì)胞、cd4+t細(xì)胞和/或b細(xì)胞生成。
因此,本發(fā)明的一個(gè)方面是提供包括這樣的序列的核酸構(gòu)建體,所述序列編碼與至少一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段可操作地連接的至少一種恒定鏈或其變體,其中所述核酸構(gòu)建體能夠引發(fā)免疫系統(tǒng),以增強(qiáng)在受試者中施用后續(xù)疫苗后的免疫接種。
在一個(gè)實(shí)施方案中,本發(fā)明提供了包括這樣的序列的核酸構(gòu)建體,所述序列編碼與至少一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段可操作地連接的至少一種恒定鏈或其變體,其中通過在受試者中施用后續(xù)疫苗,所述核酸構(gòu)建體能夠引發(fā)免疫接種。
在一個(gè)實(shí)施方案中,在本發(fā)明的核酸構(gòu)建體中包括的恒定鏈可以由其野生型序列加以改變,而不減少ii的效應(yīng)。
因此,在一個(gè)實(shí)施方案中,包括lrmk氨基酸殘基的ii-key結(jié)構(gòu)域已通過缺失或置換例如突變加以改變。
在另一個(gè)實(shí)施方案中,ii-clip結(jié)構(gòu)域的部分已通過缺失或置換例如突變加以改變。ii-clip結(jié)構(gòu)域可以具體地通過將在位置91和99上的甲硫氨酸置換為丙氨酸而改變;這令人驚訝地增加mhcii呈遞。
在另一個(gè)實(shí)施方案中,ii可以具體地通過缺失ii的前17個(gè)氨基酸而改變;這令人驚訝地增加記憶應(yīng)答。
由此可見這些改變各自可以分開或組合發(fā)生。
在一個(gè)實(shí)施方案中,當(dāng)用于引發(fā)針對(duì)病毒、微生物例如細(xì)菌或寄生蟲的疫苗的免疫應(yīng)答時(shí),在本發(fā)明的核酸構(gòu)建體中包括的恒定鏈由其野生型序列加以改變。
在另一個(gè)實(shí)施方案中,當(dāng)用于引發(fā)癌癥疫苗或針對(duì)異常生理學(xué)應(yīng)答的疫苗的免疫應(yīng)答時(shí),在本發(fā)明的核酸構(gòu)建體中包括的恒定鏈可以由或不由其野生型序列加以改變。
在另一個(gè)實(shí)施方案中,用于引發(fā)免疫應(yīng)答的核酸構(gòu)建體和用于加強(qiáng)免疫應(yīng)答的后續(xù)疫苗的至少一個(gè)部分是等同的。引發(fā)物和加強(qiáng)物(booster)的所述至少一個(gè)等同部分可以是ii或其變體、抗原肽或抗原肽的部分、或遍在的輔助t細(xì)胞表位。
同樣本發(fā)明的一個(gè)目的是提供包括如本文詳述的核酸構(gòu)建體的遞送媒介物(vehicle),其中所述遞送媒介物是基于rna的媒介物、基于dna的媒介物/載體、基于脂質(zhì)的媒介物、基于聚合物的媒介物或病毒衍生的或dna或rna媒介物。
在一個(gè)優(yōu)選實(shí)施方案中,所述遞送媒介物包括脂質(zhì)體的形成、生物可降解聚合物微球體的形成、核酸構(gòu)建體包被到膠體金顆粒上或摻入病毒衍生的dna或rna載體內(nèi)。
在另外一個(gè)優(yōu)選實(shí)施方案中,所述核酸構(gòu)建體或所述遞送媒介物借助于針注射、基因槍、射流注射(jetinjection)、電穿孔、超聲或水動(dòng)力遞送進(jìn)行施用。
因此,本發(fā)明的一個(gè)方面是提供通過包括這樣的序列的核酸構(gòu)建體刺激免疫應(yīng)答的方法,所述序列編碼與至少一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段可操作地連接的至少一種恒定鏈或其變體。
一個(gè)進(jìn)一步目的提供刺激在這些中的任何或這些中的任何的任何部分內(nèi)編碼的核酸構(gòu)建體或蛋白質(zhì)的細(xì)胞內(nèi)散布的裝置(means)。
本發(fā)明的另外一個(gè)目的是提供如由本文描述的核酸構(gòu)建體編碼的嵌合蛋白質(zhì)。
本發(fā)明的一個(gè)進(jìn)一步方面是提供識(shí)別由本文描述的核酸構(gòu)建體編碼的嵌合蛋白質(zhì)的抗體。
本發(fā)明的一個(gè)方面是提供用于改善疫苗效力的方法,其包括施用如本文詳述的核酸構(gòu)建體。
與本發(fā)明特別相關(guān)的是包括這樣的序列的核酸構(gòu)建體,所述序列編碼與至少一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段可操作地連接的至少一種恒定鏈或其變體,其適合于引發(fā)免疫應(yīng)答。
本發(fā)明的另外一個(gè)方面是提供部分試劑盒(kitinparts),所述試劑盒包括如本文描述的核酸構(gòu)建體連同施用所述核酸構(gòu)建體的醫(yī)療器械或其他裝置,和/或合適疫苗,和另外關(guān)于如何使用部分試劑盒的說明書。
因此可見本發(fā)明提供了通過給動(dòng)物施用如本文下文詳述的核酸構(gòu)建體用于加強(qiáng)動(dòng)物中的免疫應(yīng)答的方法。
本發(fā)明涉及下列內(nèi)容:
“權(quán)利要求1.一種核酸構(gòu)建體,其包括編碼下述的序列:
a.與b可操作地連接的至少一種恒定鏈或其變體,
b.至少一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段,
其中所述核酸構(gòu)建體用于癌癥疫苗的引發(fā),和/或所述恒定鏈或其變體不包括前17個(gè)氨基酸,和/或所述恒定鏈或其變體不包括key區(qū)域的lrmk氨基酸殘基,和/或所述恒定鏈包括clip區(qū)域的變體;
2.根據(jù)權(quán)利要求1的核酸構(gòu)建體,其用作用于刺激免疫應(yīng)答的引發(fā)物,從而增加后續(xù)施用的疫苗的效力;
3.根據(jù)權(quán)利要求1的核酸構(gòu)建體,其中至少一種恒定鏈或其變體是生物特異性的;
4.根據(jù)前述權(quán)利要求中任一項(xiàng)的核酸構(gòu)建體,其中至少一種恒定鏈或其變體是人源的;
5.根據(jù)前述權(quán)利要求中任一項(xiàng)的核酸構(gòu)建體,其中所述至少一種恒定鏈的至少一個(gè)區(qū)域、肽或結(jié)構(gòu)域加入、去除或置換所述至少一種恒定鏈的區(qū)域、肽或結(jié)構(gòu)域;
6.根據(jù)前述權(quán)利要求中任一項(xiàng)的核酸構(gòu)建體,其中所述至少一種恒定鏈的key區(qū)域的lrmk氨基酸殘基中的1、2、3或4個(gè)由其他氨基酸殘基置換;
7.根據(jù)前述權(quán)利要求中任一項(xiàng)的核酸構(gòu)建體,其中所述至少一種恒定鏈的key區(qū)域的lrmk氨基酸殘基中的1、2、3或4個(gè)由丙氨酸置換;
8.根據(jù)前述權(quán)利要求中任一項(xiàng)的核酸構(gòu)建體,其中所述至少一種恒定鏈的key區(qū)域的lrmk氨基酸殘基中的1、2、3或4個(gè)是缺失的;
9.根據(jù)前述權(quán)利要求中任一項(xiàng)的核酸構(gòu)建體,其中所述至少一種恒定鏈的clip區(qū)域在位置91和99上的甲硫氨酸氨基酸殘基由其他氨基酸殘基置換;
10.根據(jù)前述權(quán)利要求中任一項(xiàng)的核酸構(gòu)建體,其中所述至少一種恒定鏈的clip區(qū)域在位置91和99上的甲硫氨酸氨基酸殘基由丙氨酸置換;
11.根據(jù)前述權(quán)利要求中任一項(xiàng)的核酸構(gòu)建體,其中所述至少一種恒定鏈的前17個(gè)氨基酸是缺失的(δ17ii);
12.根據(jù)前述權(quán)利要求中任一項(xiàng)的核酸構(gòu)建體,其中所述恒定鏈包括與來自另一種蛋白質(zhì)的三聚結(jié)構(gòu)域偶聯(lián)的氨基酸殘基編號(hào)50–118;
13.根據(jù)前述權(quán)利要求中任一項(xiàng)的核酸構(gòu)建體,其中所述至少一種恒定鏈或其片段引起以cd4+t細(xì)胞依賴性和/或不依賴性方式的mhc-i和/或mhc-ii依賴性免疫應(yīng)答;
14.根據(jù)前述權(quán)利要求中任一項(xiàng)的核酸構(gòu)建體,其中至少一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段選自:致病性生物、癌癥特異性多肽和與異常生理學(xué)應(yīng)答相關(guān)的蛋白質(zhì)或肽;
15.根據(jù)前述權(quán)利要求中任一項(xiàng)的核酸構(gòu)建體,其中來自致病性生物的所述至少一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段選自包括下述的病原體:病毒、微生物和寄生蟲;
16.根據(jù)前述權(quán)利要求中任一項(xiàng)的核酸構(gòu)建體,其中至少一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段來自哺乳動(dòng)物生物,優(yōu)選人病原體;
17.根據(jù)前述權(quán)利要求中任一項(xiàng)的核酸構(gòu)建體,其中至少一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段來自癌癥特異性多肽;
18.根據(jù)前述權(quán)利要求中任一項(xiàng)的核酸構(gòu)建體,其中至少一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段來自選自下述的癌癥特異性多肽:hpv衍生的病毒癌基因e5、e6、e7和l1;存活素、bcl-xl、mcl-1和rho-c;
19.根據(jù)前述權(quán)利要求中任一項(xiàng)的核酸構(gòu)建體,其中至少一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段是與權(quán)利要求14–18所述抗原中的任何至少85%等同的抗原肽或蛋白質(zhì);
20.根據(jù)前述權(quán)利要求中任一項(xiàng)的核酸構(gòu)建體,其中在所述恒定鏈或其變體和所述抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段之間的操作接頭選自:直接連接或由間隔區(qū)介導(dǎo)的連接;
21.根據(jù)權(quán)利要求20的核酸構(gòu)建體,其中所述操作接頭是間隔區(qū);
22.根據(jù)權(quán)利要求21的核酸構(gòu)建體,其中所述間隔區(qū)編碼關(guān)于mhcii類分子的至少一個(gè)輔助表位;
23.根據(jù)權(quán)利要求21的核酸構(gòu)建體,其中所述間隔區(qū)編碼至少一個(gè)蛋白酶切割位點(diǎn);
24.根據(jù)前述權(quán)利要求中任一項(xiàng)的核酸構(gòu)建體,其中至少一種恒定鏈與至少2種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段可操作地連接;
25.根據(jù)前述權(quán)利要求中任一項(xiàng)的核酸構(gòu)建體,其中使得所述構(gòu)建體能夠表達(dá)的啟動(dòng)子在所述至少一種可操作地連接的恒定鏈或其變體和至少一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段編碼序列前;
26.根據(jù)前述權(quán)利要求中任一項(xiàng)的核酸構(gòu)建體,其中所述啟動(dòng)子選自組成型啟動(dòng)子、誘導(dǎo)型啟動(dòng)子、生物特異性啟動(dòng)子、組織特異性啟動(dòng)子和細(xì)胞類型特異性啟動(dòng)子、cmv啟動(dòng)子、sv40啟動(dòng)子和rsv啟動(dòng)子;
27.根據(jù)前述權(quán)利要求中任一項(xiàng)的核酸構(gòu)建體,其包括與抗原呈遞和/或細(xì)胞內(nèi)散布有關(guān)的基因;
28.根據(jù)前述權(quán)利要求中任一項(xiàng)的核酸構(gòu)建體,其包括編碼佐劑的基因;
29.一種遞送媒介物,其包括根據(jù)前述權(quán)利要求中任一項(xiàng)的核酸構(gòu)建體;
30.根據(jù)權(quán)利要求29的遞送媒介物,其中所述媒介物選自:基于rna的媒介物、基于dna的媒介物/載體、基于脂質(zhì)的媒介物、基于聚合物的媒介物和病毒衍生的dna或rna媒介物;
31.根據(jù)權(quán)利要求30的遞送媒介物,其中所述遞送媒介物是聚乙二醇化的載體或媒介物;
32.根據(jù)權(quán)利要求30的遞送媒介物,其中所述基于脂質(zhì)的媒介物是脂質(zhì)體;
33.根據(jù)權(quán)利要求30的遞送媒介物,其中所述基于聚合物的媒介物由選自下述的陽離子聚合物dna載體形成:聚乙烯亞胺(pei)、聚乙二胺和聚丙胺樹狀聚體、聚烯丙胺、陽離子右旋糖酐、殼聚糖、陽離子蛋白質(zhì)(聚賴氨酸、魚精蛋白和組蛋白)、陽離子肽、多肽、脂多糖和多糖;
34.根據(jù)權(quán)利要求33的遞送媒介物,其中所述遞送媒介物是生物可降解聚合物微球體;
35.根據(jù)權(quán)利要求30的遞送媒介物,其中所述遞送媒介物包括將所述核酸構(gòu)建體包被到膠體金顆粒上;
36.一種用于施用根據(jù)權(quán)利要求29–35中任一項(xiàng)的遞送媒介物的方法,其中所述施用選自針注射、基因槍、射流注射、電穿孔、超聲和水動(dòng)力遞送;
37.一種用于施用根據(jù)權(quán)利要求29–35中任一項(xiàng)的核酸構(gòu)建體的方法,其中所述施用選自針注射、基因槍、射流注射、電穿孔、超聲和水動(dòng)力遞送;
38.一種細(xì)胞,其包括如權(quán)利要求1–28中任一項(xiàng)中定義的核酸構(gòu)建體;
39.一種嵌合蛋白質(zhì),其包括至少一種可操作地連接的恒定鏈或其變體和至少一種抗原蛋白質(zhì)或肽,編碼其的序列如通過權(quán)利要求1–28中任一項(xiàng)中的核酸構(gòu)建體中的任何定義的;
40.一種抗體,其可以識(shí)別如權(quán)利要求39中定義的嵌合蛋白質(zhì);
41.根據(jù)權(quán)利要求40的抗體在用于檢測所述抗體與之結(jié)合的蛋白質(zhì)的測定中的用途;
42.一種組合物,其包括編碼下述的核酸序列:
a.與b可操作地連接的至少一種恒定鏈或其變體,
b.至少一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段;
43.一種用作藥物的組合物,其包括編碼下述的核酸序列:
a.與b可操作地連接的至少一種恒定鏈或其變體,
b.至少一種多肽;
44.根據(jù)權(quán)利要求42和43中任一項(xiàng)的組合物,其包括如權(quán)利要求1–28中任一項(xiàng)中定義的核酸構(gòu)建體;
45.根據(jù)權(quán)利要求42-44中任一項(xiàng)的組合物,其中所述組合物包括至少2種載體;
46.一種組合物,其包括根據(jù)權(quán)利要求39的至少一種嵌合蛋白質(zhì),如通過權(quán)利要求1–28中任一項(xiàng)中的核酸構(gòu)建體中的任何定義的;
47.根據(jù)權(quán)利要求42-46中任一項(xiàng)的組合物,其中所述組合物包括佐劑;
48.一種部分試劑盒,其包括:
a.包括根據(jù)權(quán)利要求1–28中任一項(xiàng)的核酸構(gòu)建體的組合物,
b.用于施用所述組合物的醫(yī)療器械或其他裝置,
c.關(guān)于如何使用所述部分試劑盒的說明書;
49.根據(jù)權(quán)利要求48的部分試劑盒,其中所述試劑盒包括第二種活性成分;
50.根據(jù)權(quán)利要求49的部分試劑盒,其中所述第二種活性成分是合適疫苗;
51.一種用于增加疫苗效力的方法,其包括步驟:
a.提供根據(jù)權(quán)利要求1–28中任一項(xiàng)的核酸構(gòu)建體、或根據(jù)權(quán)利要求42–47中任一項(xiàng)的組合物,
b.通過施用步驟a)的核酸構(gòu)建體或組合物引發(fā)受試者的免疫系統(tǒng),從而刺激所述受試者中的免疫應(yīng)答,和
c.通過施用合適疫苗加強(qiáng)步驟b)的免疫應(yīng)答;
52.根據(jù)權(quán)利要求51的方法,其中用于引發(fā)免疫應(yīng)答的所述核酸構(gòu)建體的至少一部分和用于加強(qiáng)所述免疫應(yīng)答的所述疫苗是等同的;
53.根據(jù)權(quán)利要求52的方法,其中所述等同部分是所述抗原肽或蛋白質(zhì)的部分;
54.根據(jù)權(quán)利要求52的方法,其中所述等同部分是遍在輔助t細(xì)胞表位;
55.根據(jù)權(quán)利要求51的方法,其中所述免疫應(yīng)答是mhc-i依賴性的;
56.根據(jù)權(quán)利要求51的方法,其中所述免疫應(yīng)答是mhc-ii依賴性的;
57.根據(jù)權(quán)利要求51的方法,其中所述免疫應(yīng)答是mhc-i和mhc-ii依賴性的;
58.根據(jù)權(quán)利要求51的方法,其中所述免疫應(yīng)答是t細(xì)胞依賴性的;
59.根據(jù)權(quán)利要求51的方法,其中所述免疫應(yīng)答是cd4+t細(xì)胞依賴性的;
60.根據(jù)權(quán)利要求51的方法,其中所述免疫應(yīng)答是cd4+t細(xì)胞不依賴性的;
61.根據(jù)權(quán)利要求51的方法,其中所述免疫應(yīng)答是cd8+t細(xì)胞依賴性的;
62.根據(jù)權(quán)利要求51–61中任一項(xiàng)的方法,其包括步驟:
a.提供根據(jù)權(quán)利要求42–47中任一項(xiàng)的至少一種組合物,
b.給受試者施用所述至少一種組合物至少一次用于動(dòng)物的治療或預(yù)防;
63.根據(jù)權(quán)利要求51–62中任一項(xiàng)的方法,其中所述組合物施用于最可能是給定感染的接受器的區(qū)域;
64.根據(jù)權(quán)利要求51–63中任一項(xiàng)的方法,其中所述受試者是人;
65.根據(jù)權(quán)利要求51–63中任一項(xiàng)的方法,其中所述受試者是哺乳動(dòng)物;
66.根據(jù)權(quán)利要求51–63中任一項(xiàng)的方法,其中所述受試者是雪貂;
67.根據(jù)權(quán)利要求51–63中任一項(xiàng)的方法,其中所述受試者是魚;
68.根據(jù)權(quán)利要求51–63中任一項(xiàng)的方法,其中所述受試者是鳥。”。
附圖說明
圖1:用基于ii鏈的裸露dna疫苗的dna引發(fā)。
圖2:在ii序列中結(jié)構(gòu)域和測試的突變的定位。
圖3:ii顯著增加siinfekl/h-2kbova衍生表位的細(xì)胞表面呈遞。
圖4:ii僅以順式工作。
圖5:n末端缺失和置換不影響ii刺激能力。
圖6:c末端缺失和置換不影響ii刺激能力。
圖7:僅n和c末端缺失降低ii刺激能力。
圖8:ad-iigp和ad-gp疫苗的劑量應(yīng)答。
圖9:ad-gp、ad-iigp和ad-iiclipgp作用于mhcii類呈遞的比較。
圖10:ad-gp、ad-iigp、ad-gplamp-1和ad-iiδ17gp在體內(nèi)時(shí)間過程研究中的比較。
圖11:ad-gp、ad-iigp、ad-iiδ17gp、ad-iikeygp、ad-iiclipgp、ad-ii1-117gp和ad-ii1-199gp在體內(nèi)應(yīng)答中的比較。
圖12:ad-gp能夠引發(fā)后續(xù)ad-iigp加強(qiáng)。
圖13:ad-iigp不能引發(fā)后續(xù)ad-gp或ad-iigp加強(qiáng)。
圖14:ad-gp和adii-gp疫苗的劑量應(yīng)答。
圖15:與包括殘基50-215的恒定鏈變體(ii50-215)偶聯(lián)的甘露糖受體,與腺病毒纖維蛋白質(zhì)進(jìn)一步偶聯(lián)。
定義
腺病毒:一組包含雙鏈dna的病毒。腺病毒可以進(jìn)行遺傳修飾,使得其不能復(fù)制或條件性不能復(fù)制。以這種形式,作為腺病毒構(gòu)建體或腺病毒載體,它們可以用作基因遞送媒介物用于疫苗接種或基因療法。
腺病毒纖維蛋白質(zhì):來自腺病毒的任何血清型的纖維蛋白質(zhì)。它還稱為腺病毒纖維結(jié)或具有異源結(jié)插入片段的腺病毒纖維結(jié)。
佐劑:其與施用的免疫原決定簇/抗原/核酸構(gòu)建體的混合物增加或以其他方式修飾對(duì)于所述決定簇的免疫應(yīng)答的任何物質(zhì)。
氨基酸:任何合成或天然存在的氨基羧酸,包括在肽和多肽包括在體內(nèi)合成的蛋白質(zhì)和酶中存在的任何氨基酸,從而包括氨基酸的修飾。術(shù)語氨基酸在本文中與術(shù)語“氨基酸殘基”同義使用,其意欲包含如所述已與至少一個(gè)其他種類例如2、例如3、例如超過3個(gè)其他種類反應(yīng)的氨基酸。通用詞氨基酸包括天然和非天然氨基酸,其中任何可以以“d”或“l(fā)”異構(gòu)形式存在。氨基酸可以縮寫為‘a(chǎn)a’。
抗體:免疫球蛋白分子和免疫球蛋白分子的活性部分。抗體是例如完整免疫球蛋白分子或其保留免疫學(xué)活性的片段。
抗原:可以與克隆分布的免疫受體(t細(xì)胞或b細(xì)胞受體)結(jié)合的任何物質(zhì)。通常是肽、多肽或多聚體多肽。抗原優(yōu)選能夠引發(fā)免疫應(yīng)答。
加強(qiáng):通過加強(qiáng)物注射(shot)或劑量來加強(qiáng)是給予免疫試劑例如疫苗的一次或多次另外劑量,在用于引發(fā)免疫系統(tǒng)的物質(zhì)的初始劑量后一段時(shí)間給予,以維持或增強(qiáng)通過相同(同源)或另一種(異源)免疫試劑的先前劑量引發(fā)的免疫應(yīng)答。
載體:抗原與之偶聯(lián)以幫助誘導(dǎo)免疫應(yīng)答的實(shí)體或化合物。
嵌合蛋白質(zhì):由核苷酸序列編碼的遺傳改造蛋白質(zhì),所述核苷酸序列通過將2種或更多種完整或部分基因或一系列(非)隨機(jī)核酸剪接在一起制備。
補(bǔ)體:其作用于“補(bǔ)充”抗體工作的復(fù)雜系列的血液蛋白質(zhì)。補(bǔ)體破壞細(xì)菌,產(chǎn)生炎癥,且調(diào)節(jié)免疫反應(yīng)。
細(xì)胞因子:生長或分化調(diào)節(jié)劑,在本文中非限定使用,并且不應(yīng)限制本發(fā)明和權(quán)利要求的解釋。除細(xì)胞因子外,粘附或輔助分子或其任何組合可以單獨(dú)或與細(xì)胞因子組合采用。
ctl:細(xì)胞毒性t淋巴細(xì)胞。表達(dá)cd8連同t細(xì)胞受體,且因此能夠響應(yīng)由i類分子呈遞的抗原的t細(xì)胞亞組。
遞送媒介物:由此核苷酸序列或多肽或兩者可以從至少一種介質(zhì)轉(zhuǎn)運(yùn)到另一種介質(zhì)的實(shí)體。
片段:用于指出核酸或多肽的非全長部分。因此,片段自身也分別是核酸或多肽。
異源加強(qiáng)或引發(fā)-加強(qiáng):其中用于加強(qiáng)免疫系統(tǒng)的物質(zhì)不同于先前用于引發(fā)免疫應(yīng)答的物質(zhì)。
同源加強(qiáng)或引發(fā)-加強(qiáng):其中用于加強(qiáng)免疫系統(tǒng)的物質(zhì)與先前用于引發(fā)免疫應(yīng)答的物質(zhì)相同。
個(gè)體:鳥、哺乳動(dòng)物、魚、兩棲類或爬行類包括人類的任何物種或亞種。
恒定鏈:與內(nèi)質(zhì)網(wǎng)和后續(xù)細(xì)胞區(qū)室中的mhcii分子結(jié)合且穩(wěn)定其的整合膜蛋白質(zhì)糖蛋白。此處術(shù)語恒定鏈涵蓋所有天然存在或人工生成的與人恒定鏈具有特定相似性的全長或片段化的同源基因和蛋白質(zhì)。恒定鏈在本文中縮寫為ii。
分離的:與本文公開的核酸、多肽或抗體結(jié)合使用的‘分離的’指已鑒定且從其天然的、一般的細(xì)胞環(huán)境的組分中分離和/或回收的那些。本發(fā)明的核酸、多肽和抗體優(yōu)選是分離的,并且本發(fā)明的疫苗和其他組合物優(yōu)選包括分離核酸、多肽或分離抗體。
mhc:主要組織相容性復(fù)合物,存在mhc的2個(gè)主要亞類,i類和ii類。
裸露dna:不與組蛋白結(jié)合的dna;通常是低甲基化和富含cpg的dna。裸露dna可以是環(huán)狀或線性的,例如環(huán)狀質(zhì)粒。
核酸:傳送遺傳信息的核苷酸鏈或序列。就本發(fā)明而言,核酸可以是脫氧核糖核酸(dna)或下述中的任何:核糖核酸(rna)、鎖核酸(lna)、肽核酸(pna)、嵌入核酸(ina)、扭曲嵌入核酸(tina)、己糖醇核酸(hna)、阿糖核酸(ana)、環(huán)己烷核酸(cna)、環(huán)己烯基核酸(cena)、甘油核酸(gna)、蘇糖基核酸(tna)、缺口-聚體(gap-mers)、混合-聚體(mix-mers)和嗎啉代類物質(zhì)(morpholinos)。
核酸構(gòu)建體:遺傳改造的核酸。一般包括幾種元件例如基因或其片段、啟動(dòng)子、增強(qiáng)子、終止子、多聚腺苷酸尾、接頭、多聚接頭、操作接頭、多克隆位點(diǎn)(mcs)、標(biāo)記、終止密碼子、其他調(diào)節(jié)元件、內(nèi)部核糖體進(jìn)入位點(diǎn)(ires)或其他。
操作接頭:以保證核酸或多肽的生物加工的方式使核酸構(gòu)建體或(嵌合)多肽的2個(gè)部分結(jié)合在一起的核苷酸或氨基酸殘基序列。
病原體:疾病的特異性成因劑,特別是生物劑例如病毒、細(xì)菌、朊病毒或寄生蟲,其可以引起對(duì)其宿主的疾病,也被稱為傳染劑。
肽:限定序列且由酰胺鍵連接的多個(gè)共價(jià)連接的氨基酸殘基。該術(shù)語與寡肽和多肽類似地使用。天然和/或非天然氨基酸可以通過肽鍵或通過非肽鍵連接。術(shù)語肽還包含通過化學(xué)或酶催化反應(yīng)引入的翻譯后修飾,如本領(lǐng)域已知的。該術(shù)語可以指多肽變體或片段。
藥物載體:也稱為賦形劑或穩(wěn)定劑,在采用的劑量和濃度下對(duì)暴露于其的細(xì)胞或個(gè)體是無毒的。通常生理學(xué)上可接受的載體是ph緩沖的水性溶液。生理學(xué)上可接受的載體的例子包括緩沖液例如磷酸鹽、檸檬酸鹽和其他有機(jī)酸;抗氧化劑包括抗壞血酸;低分子量(小于約10個(gè)殘基)多肽;蛋白質(zhì)例如血清白蛋白、明膠或免疫球蛋白;親水聚合物例如聚乙烯吡咯烷酮;氨基酸例如甘氨酸、谷氨酰胺、天冬酰胺、精氨酸或賴氨酸;單糖、二糖和其他碳水化合物包括葡萄糖、甘露糖或糊精;螯合劑例如edta;糖醇例如甘露糖醇或山梨糖醇;成鹽抗衡離子例如鈉;和/或非離子表面活性劑例如tweentm、聚乙二醇(peg)和pluronicstm。
多個(gè):至少2個(gè)。
引發(fā):用例如dna初始引發(fā)免疫系統(tǒng),以使免疫應(yīng)答集中于所需免疫原。
引發(fā)-加強(qiáng):用例如dna初始引發(fā)免疫系統(tǒng),以使免疫應(yīng)答集中于所需免疫原,并且用疫苗隨后加強(qiáng)免疫應(yīng)答,導(dǎo)致由所述疫苗誘導(dǎo)的免疫應(yīng)答中的增加。
啟動(dòng)子:rna聚合酶在其上結(jié)合以起始通過一種或多種附近結(jié)構(gòu)基因的信使rna轉(zhuǎn)錄的dna鏈中的結(jié)合位點(diǎn)。
信號(hào)肽:決定蛋白質(zhì)在細(xì)胞中的最終定位的短氨基酸序列,也被稱為分選肽。
合適疫苗:用于根據(jù)本發(fā)明使用的任何疫苗,能夠加強(qiáng)通過初始引發(fā)刺激的免疫應(yīng)答,特征在于用于引發(fā)免疫應(yīng)答的核酸構(gòu)建體和用于加強(qiáng)免疫應(yīng)答的后續(xù)疫苗的至少一個(gè)部分是等同的。引發(fā)物和加強(qiáng)物的所述至少一個(gè)等同部分可以是ii或其變體、抗原肽或抗原肽的部分、或遍在的輔助t細(xì)胞表位。
表面活性劑:能夠減少它溶解于其中的液體的表面張力的表面活性試劑。表面活性劑是包含其為親水的極性基團(tuán)和其為疏水的非極性基團(tuán),并且通常由脂肪鏈組成的化合物。
疫苗:能夠在動(dòng)物中誘導(dǎo)免疫應(yīng)答的物質(zhì)或組合物:在該正文中也被稱為免疫原性組合物。免疫應(yīng)答是在生物中誘導(dǎo)記憶的免疫應(yīng)答(體液/抗體和/或細(xì)胞免疫應(yīng)答),導(dǎo)致傳染劑由二次而不是初次應(yīng)答對(duì)抗,從而減少其對(duì)宿主生物的影響。本發(fā)明的疫苗可以作為預(yù)防和/或治療藥物給予。組合物可以包括下述中的一種或多種:一種或多種抗原、包括與ii可操作地連接的一種或多種抗原的核酸構(gòu)建體、載體、佐劑和藥物載體。
變體:給定參考核酸或多肽的‘變體’指這樣的核酸或多肽,其展示出與所述參考核酸或多肽的一定序列同源性/同一性程度,但不等同于所述參考核酸或多肽。
結(jié)構(gòu)域、區(qū)域和基序在本文中可以互換使用。
具體實(shí)施方式
本發(fā)明涉及包括這樣的序列的核酸構(gòu)建體,所述序列編碼與至少一種抗原蛋白質(zhì)或肽編碼序列可操作地連接的恒定鏈或其變體。核酸構(gòu)建體用于引發(fā)免疫應(yīng)答,以加強(qiáng)后續(xù)加強(qiáng)疫苗接種的效應(yīng)。
免疫應(yīng)答
疫苗可以在預(yù)防上使用:它們?cè)趯?shí)際感染發(fā)生前給予;或在治療上使用:其中它們引發(fā)或加速對(duì)已在體內(nèi)的病原體的免疫應(yīng)答。2種疫苗接種方法要求實(shí)體免疫應(yīng)答的建立。通過感染或疫苗接種激活的免疫應(yīng)答依賴于幾種細(xì)胞類型例如t、b和抗原呈遞細(xì)胞,以及幾種不同分子主要是抗原、mhc分子、t和b細(xì)胞受體及許多其他的相互作用。
抗原是在抗原呈遞細(xì)胞的表面上由mhc分子呈遞的肽片段。抗原可以是外源的,即致病性來源,或源于生物自身,所謂的自體或自身抗原。mhc分子是由特異性染色體區(qū)域編碼的多態(tài)基因家族的代表,所述特異性染色體區(qū)域稱為“主要組織相容性復(fù)合物”(“majorhistocompatibilitycomplex”),因此為mhc。存在2類mhc分子,mhci類(mhc-i)和mhcii類(mhc-ii)。
t輔助細(xì)胞通過由mhcii類(mhc-ii)分子呈遞的抗原刺激,所述mhc-ii分子位于抗原呈遞細(xì)胞的表面上。mhc-ii分子在內(nèi)質(zhì)網(wǎng)中合成。在合成過程中,它們以阻止mhc-ii分子負(fù)載有自體或自身抗原的方式與恒定鏈(ii)組合。mhc-ii分子通過恒定鏈中的信號(hào)序列轉(zhuǎn)運(yùn)至在特異性細(xì)胞區(qū)室中的細(xì)胞表面。隨著區(qū)室通過加工其內(nèi)含物而成熟,它從溶酶體發(fā)展到晚期內(nèi)體(在與內(nèi)吞囊泡融合后)到mhcii類區(qū)室(miic)。內(nèi)吞囊泡包含外源抗原,即蛋白酶解切割的細(xì)菌肽片段。這些片段通過其降解制備,以負(fù)載到mhc-ii分子上。mhc-ii分子通過恒定鏈在2部分過程中釋放,這時(shí)恒定鏈?zhǔn)紫韧ㄟ^蛋白酶解被降解,僅留下在mhc-ii結(jié)合結(jié)構(gòu)域中稱為clip的肽,其次通過hla-dm分子去除clip。mhc-ii分子隨后游離,以結(jié)合外源抗原,且在miic囊泡與質(zhì)膜融合后,將其呈遞在細(xì)胞表面上。這起始體液免疫應(yīng)答,因?yàn)槌蔬f的抗原刺激t輔助細(xì)胞的激活,這依次通過幾種方式激活b細(xì)胞,這最終分化成抗體分泌細(xì)胞。
當(dāng)t細(xì)胞毒性細(xì)胞的t細(xì)胞受體識(shí)別與在抗原呈遞細(xì)胞上的mhci類分子結(jié)合的抗原時(shí),細(xì)胞免疫應(yīng)答起始。mhc-i分子不與功能性類似與mhc-ii結(jié)合的恒定鏈的分子結(jié)合。mhc-i加工成的抗原呈遞分子另外不同于mhc-ii分子的那種,因?yàn)閙hc-i分子已在內(nèi)質(zhì)網(wǎng)中負(fù)載有抗原。由mhc-i分子呈遞的抗原一般是通過蛋白質(zhì)的蛋白酶體切割的肽片段,所述蛋白質(zhì)已通過抗原呈遞細(xì)胞自身合成。這些蛋白質(zhì)可以是在細(xì)胞自身dna中編碼的異常蛋白質(zhì)或衍生自病毒或其他病原體的蛋白質(zhì),所述病毒或其他病原體已感染細(xì)胞且寄生其蛋白質(zhì)合成機(jī)制。mhci類相關(guān)蛋白酶解系統(tǒng)存在于基本上所有細(xì)胞中。
2類t細(xì)胞的功能是顯著不同的,如由其名稱所示的。細(xì)胞毒性t細(xì)胞通過細(xì)胞的直接裂解和通過分泌細(xì)胞因子例如γ-干擾素來消滅細(xì)胞內(nèi)病原體和腫瘤。占優(yōu)勢的細(xì)胞毒性t細(xì)胞是cd8+t細(xì)胞,這也是抗原特異性的。輔助t細(xì)胞也是裂解細(xì)胞,但其主要功能是分泌促進(jìn)b細(xì)胞(抗體生產(chǎn)細(xì)胞)和其他t細(xì)胞的活性的細(xì)胞因子,并且因此它們廣泛增強(qiáng)對(duì)于外源抗原的免疫應(yīng)答,包括抗體介導(dǎo)的和細(xì)胞毒性t細(xì)胞介導(dǎo)的應(yīng)答機(jī)制。cd4+t細(xì)胞是免疫應(yīng)答中的主要輔助t細(xì)胞表型。
核酸構(gòu)建體
本發(fā)明的一個(gè)方面涉及包括這樣的序列的核酸構(gòu)建體例如裸露dna構(gòu)建體,所述序列編碼與至少一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段簡言之抗原可操作地連接的至少一種恒定鏈或其變體。
在一個(gè)實(shí)施方案中,本發(fā)明涉及包括這樣的序列的核酸構(gòu)建體,所述序列編碼與至少一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段可操作地連接的至少一種恒定鏈或其變體,其中所述恒定鏈或其變體不包括key區(qū)域的lrmk氨基酸殘基。
在另一個(gè)實(shí)施方案中,本發(fā)明涉及包括這樣的序列的核酸構(gòu)建體,所述序列編碼與至少一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段可操作地連接的至少一種恒定鏈或其變體,其中所述恒定鏈或其變體包括clip區(qū)域的變體。
在另一個(gè)實(shí)施方案中,本發(fā)明涉及包括這樣的序列的核酸構(gòu)建體,所述序列編碼與至少一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段可操作地連接的至少一種恒定鏈或其變體,其中所述恒定鏈或其變體不包括前17個(gè)氨基酸。
在另外一個(gè)實(shí)施方案中,本發(fā)明涉及包括這樣的序列的核酸構(gòu)建體,所述序列編碼與至少一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段可操作地連接的至少一種恒定鏈或其變體,其中所述核酸構(gòu)建體用于癌癥疫苗的引發(fā)。
核酸構(gòu)建體應(yīng)理解為遺傳改造的核酸。核酸構(gòu)建體可以是非復(fù)制的且線性的核酸、環(huán)狀表達(dá)載體或自主復(fù)制質(zhì)粒。核酸構(gòu)建體可以包括幾種元件,例如但不限于基因或其片段、啟動(dòng)子、增強(qiáng)子、終止子、多聚腺苷酸尾、接頭、多聚接頭、操作接頭、多克隆位點(diǎn)(mcs)、標(biāo)記、終止密碼子、內(nèi)部核糖體進(jìn)入位點(diǎn)(ires)和用于整合的宿主同源序列或其他限定元件。應(yīng)當(dāng)理解根據(jù)本發(fā)明的核酸構(gòu)建體可以包括上述元件的任何組合的所有或亞組。
用于改造核酸構(gòu)建體的方法是本領(lǐng)域眾所周知的(參見例如,molecularcloning:alaboratorymanual,sambrook等人,編輯,coldspringharborlaboratory,第2版,coldspringharbor,n.y.,1989)。進(jìn)一步地,根據(jù)本發(fā)明的核酸構(gòu)建體可以無需模板而合成,并且可以得自各種商業(yè)供應(yīng)者(例如genscriptcorporation)。
在一個(gè)實(shí)施方案中,核酸構(gòu)建體包括的核酸殘基可以被修飾。所述修飾可以選自:乙酰化、甲基化、磷酸化、遍在蛋白化、核糖基化、硫化及其他。
在一個(gè)實(shí)施方案中,根據(jù)本發(fā)明的核酸構(gòu)建體可以由dna組成。在另一個(gè)實(shí)施方案中,核酸構(gòu)建體可以由選自下述的核酸組成:脫氧核糖核酸(dna)、核糖核酸(rna)、鎖核酸(lna)、肽核酸(pna)、嵌入核酸(ina)、扭曲嵌入核酸(tina)、己糖醇核酸(hna)、阿糖核酸(ana)、環(huán)己烷核酸(cna)、環(huán)己烯基核酸(cena)、甘油核酸(gna)、蘇糖基核酸(tna)、缺口-聚體、混合-聚體、嗎啉代類物質(zhì)或其組合。
如上所述,mhcii分子在加工過程中與恒定鏈結(jié)合,直至結(jié)合的恒定鏈降解,以允許外源抗原肽負(fù)載到mhcii分子上。當(dāng)應(yīng)用包括恒定鏈–接頭–表位的‘外部’蛋白質(zhì)構(gòu)建體時(shí),其中恒定鏈包括key區(qū)域(包括lrmk氨基酸殘基),所述蛋白質(zhì)構(gòu)建體在所述mhcii分子定位于細(xì)胞的細(xì)胞外表面上時(shí)將與mhcii分子(包含抗原)相互作用。因此,效應(yīng)是外部的,并且依賴于蛋白質(zhì)構(gòu)建體(包括lrmk氨基酸殘基)的恒定鏈key-殘基競爭負(fù)載到mhcii分子上的能力。換言之,在細(xì)胞內(nèi)加工過程中已負(fù)載到mhcii分子上的抗原必須被“倒出”(“tippedoff”)或從細(xì)胞表面上的mhcii分子中去除,并且由包括恒定鏈–接頭–表位的蛋白質(zhì)構(gòu)建體代替。這給出無法擴(kuò)增的“1:1”-效應(yīng),并且它不變地依賴于來自恒定鏈的lrmk氨基酸殘基的存在。
根據(jù)本發(fā)明的核酸構(gòu)建體涉及應(yīng)用編碼恒定鏈或其變體且還編碼抗原肽或表位的‘內(nèi)部’核酸構(gòu)建體,即核酸構(gòu)建體轉(zhuǎn)染到受試者中細(xì)胞的細(xì)胞內(nèi)間隙內(nèi)。所述核酸構(gòu)建體將使用細(xì)胞翻譯機(jī)制,以產(chǎn)生恒定鏈–接頭–表位或恒定鏈–表位產(chǎn)物,在所述mhc分子定位在細(xì)胞內(nèi)例如在胞內(nèi)體或miic中時(shí),這將與mhcii分子或mhci分子(不包含抗原)相互作用。因此,效應(yīng)是內(nèi)部的,并且不依賴于蛋白質(zhì)構(gòu)建體(包括lrmk氨基酸殘基)的恒定鏈key-結(jié)構(gòu)域競爭負(fù)載到mhc分子上的能力。事實(shí)上,該效應(yīng)不依賴于ii-key區(qū)域及其lrmk殘基的存在。此外,這給出可以擴(kuò)增的效應(yīng),因?yàn)橐粋€(gè)核酸構(gòu)建體可以產(chǎn)生超過一種產(chǎn)物。
核酸構(gòu)建體的‘內(nèi)部’使用具有優(yōu)于蛋白質(zhì)構(gòu)建體的‘外部’使用的幾個(gè)優(yōu)點(diǎn),如上詳述:1)擴(kuò)增;小量核酸構(gòu)建體的引入產(chǎn)生可以都與mhc分子結(jié)合的許多產(chǎn)物,這還可以是二次擴(kuò)增的,因?yàn)榕cmhc結(jié)合的所述產(chǎn)物可以通過內(nèi)在化進(jìn)一步再循環(huán),以最終增加其展示,2)細(xì)胞自身抗原加工系統(tǒng)的使用確保表位與mhc分子的正確和緊密結(jié)合,3)不需要ii-key區(qū)域的lrmk殘基和天然ii-clip結(jié)構(gòu)域。
密碼子最佳化和簡并核酸序列
功能蛋白質(zhì)在異源宿主中的表達(dá)是現(xiàn)代生物技術(shù)的基石。不幸的是,許多蛋白質(zhì)難以在其原始背景外表達(dá)。它們可以包含表達(dá)限制性調(diào)節(jié)元件,來自使用非經(jīng)典核苷酸編碼的生物,或來自具有在所需宿主中很少使用的密碼子的普遍基因(generife)。基因合成速度和效率中的改善已致使可行的完整基因重新設(shè)計(jì)用于最大限度的蛋白質(zhì)表達(dá)。例如,當(dāng)在研究下的基因的密碼子頻率與宿主表達(dá)系統(tǒng)的密碼子頻率匹配時(shí),蛋白質(zhì)表達(dá)可以顯著改善。例如,重新設(shè)計(jì)策略可以不僅包括最佳密碼子偏好的使用,還包括mrna結(jié)構(gòu)元件的改變以及翻譯和起始區(qū)的修飾。用于密碼子最佳化的技術(shù)是本領(lǐng)域技術(shù)人員已知的,并且可以通過商業(yè)供應(yīng)者例如genscriptcorporation執(zhí)行。
應(yīng)當(dāng)理解根據(jù)本發(fā)明包括恒定鏈或其變體的核酸構(gòu)建體可以以任何方式進(jìn)行密碼子最優(yōu)化,以便通過翻譯成蛋白質(zhì)即氨基酸產(chǎn)生包括根據(jù)本發(fā)明的恒定鏈或其變體的氨基酸序列,所述恒定鏈對(duì)應(yīng)于seqidno:2(人ii)中公開的氨基酸序列。
同樣地,根據(jù)本發(fā)明包括恒定鏈的核酸構(gòu)建體可以以任何方式進(jìn)行密碼子最優(yōu)化,以便通過翻譯成蛋白質(zhì)即氨基酸產(chǎn)生包括根據(jù)本發(fā)明的恒定鏈或其變體的氨基酸序列,所述恒定鏈對(duì)應(yīng)于其中核酸構(gòu)建體可以用于引發(fā)免疫應(yīng)答的任何動(dòng)物的氨基酸序列;包括任何脊椎動(dòng)物、哺乳動(dòng)物、魚或鳥。
密碼子偏好:密碼子偏好已鑒定為在原核生物基因表達(dá)中的單個(gè)最重要因子。給定密碼子在遺傳密碼中出現(xiàn)的程度在生物之間、在以高和低水平表達(dá)的蛋白質(zhì)之間和甚至在相同操縱子的不同部分之間顯著不同。關(guān)于這點(diǎn)的原因幾乎是必然的,因?yàn)閮?yōu)選密碼子與細(xì)胞內(nèi)可用的同族trna豐度相關(guān)。這個(gè)關(guān)系用于最優(yōu)化翻譯系統(tǒng)且平衡密碼子濃度與同工trna(isoacceptortrna)濃度。
替換很少使用的密碼子:一般而言,基因包含的罕見密碼子越多,異源蛋白質(zhì)將在那種特定宿主系統(tǒng)內(nèi)以合理水平表達(dá)的可能性越小。如果罕見密碼子成簇或在蛋白質(zhì)的n末端部分中出現(xiàn),那么這些水平變得甚至更低。用更緊密地反映宿主系統(tǒng)的密碼子偏好而不改變氨基酸序列的其他密碼子替換罕見密碼子可以增加功能蛋白質(zhì)表達(dá)水平。
消除問題密碼子:生物使用小于5%-10%時(shí)機(jī)的任何密碼子都可能引起問題,不管它來自何處。再次,緊密或相鄰密碼子對(duì)蛋白質(zhì)表達(dá)比它們分開可以具有更大的影響。消除罕見密碼子和可以作為終止信號(hào)閱讀的密碼子可以阻止低表達(dá)或不存在表達(dá)的情況。
在哺乳動(dòng)物宿主中表達(dá)病毒蛋白質(zhì):如果基因正確制備,甚至病毒基因也可以在哺乳動(dòng)物細(xì)胞系中成功表達(dá)。病毒基因的密集信息負(fù)荷頻繁導(dǎo)致重疊的讀碼框。許多病毒基因還在編碼序列內(nèi)編碼順式作用的負(fù)調(diào)節(jié)序列。病毒基因可以重新合成,不僅只表達(dá)所需蛋白質(zhì),還破壞調(diào)節(jié)元件,從而增強(qiáng)蛋白質(zhì)產(chǎn)生。病毒密碼子最優(yōu)化在dna疫苗研究中是特別有用的,因?yàn)樗黾影械拿庖咴浴?/p>
其他限制:盡管密碼子偏好在基因表達(dá)中發(fā)揮很大作用,但表達(dá)載體和轉(zhuǎn)錄啟動(dòng)子的選擇也是重要的。在蛋白質(zhì)n末端區(qū)域周圍的核苷酸序列對(duì)于罕見密碼子的存在和緊鄰起始aug的密碼子的特性是特別敏感的。在翻譯和mrna穩(wěn)定性之間還存在一定相互影響。
遺傳密碼的簡并性
根據(jù)上文可見遺傳密碼具有冗余性但不具有多義性(ambiguity)。例如,盡管密碼子gaa和gag都指定谷氨酸(冗余性),但其中無一指定任何其他氨基酸(無多義性)(關(guān)于完全關(guān)聯(lián)參見下文密碼子表)。編碼一種氨基酸的密碼子可以在其3個(gè)位置中的任何中不同。遺傳密碼的簡并性是解釋沉默突變的存在的原因。因?yàn)?種堿基的三聯(lián)體密碼指定20種氨基酸和終止密碼子,所以產(chǎn)生簡并性。
該表顯示20種氨基酸、起始(start)和終止(stop)密碼子和64種可能密碼子。mrna的方向是5'到3'。
同義置換
沉默突變或置換是不導(dǎo)致對(duì)蛋白質(zhì)的氨基酸序列的改變的dna突變。它們可以在非編碼區(qū)中出現(xiàn)(在基因外或在內(nèi)含子內(nèi)),或它們可以以不改變最終氨基酸序列的方式在外顯子內(nèi)出現(xiàn)。短語沉默突變或置換通常可與短語同義突變或置換互換使用;然而,同義突變或置換是前者的子范疇,僅在外顯子內(nèi)出現(xiàn)。
應(yīng)當(dāng)理解根據(jù)本發(fā)明包括恒定鏈或其變體的核酸構(gòu)建體可以包括同義置換,以便通過翻譯成蛋白質(zhì)即氨基酸產(chǎn)生包括根據(jù)本發(fā)明的恒定鏈或其變體的氨基酸序列,所述恒定鏈對(duì)應(yīng)于seqidno:2(人ii)中公開的氨基酸序列。
同樣地,根據(jù)本發(fā)明包括恒定鏈的核酸構(gòu)建體可以包括同義置換,以便通過翻譯成蛋白質(zhì)即氨基酸產(chǎn)生包括根據(jù)本發(fā)明的恒定鏈或其變體的氨基酸序列,所述恒定鏈對(duì)應(yīng)于其中核酸構(gòu)建體可以用于引發(fā)免疫應(yīng)答的任何動(dòng)物的氨基酸序列;包括任何脊椎動(dòng)物、哺乳動(dòng)物、魚或鳥。
非同義置換成同義氨基酸
非同義置換引起氨基酸中的改變。然而,氨基酸根據(jù)所述氨基酸的性質(zhì)分組,并且一個(gè)氨基酸由另一個(gè)氨基酸的置換可以對(duì)包括所述氨基酸的蛋白質(zhì)的功能或性質(zhì)沒有影響(如果置換導(dǎo)致同義氨基酸)。此類置換可以指示保守置換或突變:dna或rna序列中的改變導(dǎo)致一個(gè)氨基酸由生物化學(xué)相似的氨基酸替換。
因此,應(yīng)當(dāng)理解根據(jù)本發(fā)明包括恒定鏈或其變體的核酸構(gòu)建體可以包括非同義置換,以便通過翻譯成蛋白質(zhì)即氨基酸產(chǎn)生包括恒定鏈變體的氨基酸序列,其中所述非同義置換導(dǎo)致其為同義的一個(gè)或多個(gè)氨基酸的置換。
同義置換可以包括疏水性氨基酸由另一個(gè)疏水性氨基酸的置換;親水性氨基酸由另一個(gè)親水性氨基酸的置換;極性氨基酸由另一個(gè)極性氨基酸的置換;非極性氨基酸由另一個(gè)非極性氨基酸的置換;帶正電的氨基酸由另一個(gè)帶正電的氨基酸的置換;帶負(fù)電的氨基酸由另一個(gè)帶負(fù)電的氨基酸的置換;中性氨基酸由另一個(gè)中性氨基酸的置換;多重性(ambiguous)氨基酸由其配對(duì)多重性荷電氨基酸的置換,例如異亮氨酸和亮氨酸、天冬酰胺和天冬氨酸以及谷氨酰胺和谷氨酸;芳香族氨基酸由另一個(gè)芳香族氨基酸的置換;脂肪族氨基酸由另一個(gè)脂肪族氨基酸的置換;或任何氨基酸由丙氨酸的置換。這些置換可以指示等值置換。
剪接變體
可變剪接是rna剪接變異機(jī)制,其中初級(jí)基因轉(zhuǎn)錄物(前mrna)的外顯子分離且重新連接,以便產(chǎn)生可變核糖核苷酸排列。這些線性組合隨后經(jīng)歷翻譯過程,其中指定氨基酸的特定和獨(dú)特序列,導(dǎo)致同種型蛋白質(zhì)或剪接變體。以這種方式,可變剪接使用遺傳表達(dá)以促進(jìn)更多樣的蛋白質(zhì)的合成。在真核生物中,可變剪接是朝向更高效率的重要步驟,因?yàn)樾畔⒖梢越?jīng)濟(jì)得多地被貯存。幾種蛋白質(zhì)可以在其長度在原核生物編碼方式中只夠2種蛋白質(zhì)的dna序列中編碼。
在一個(gè)實(shí)施方案中,本發(fā)明的核酸構(gòu)建體可以這樣設(shè)計(jì),以便產(chǎn)生多重抗原肽或抗原肽的片段和/或多重恒定鏈或其變體。
在一個(gè)實(shí)施方案中,根據(jù)本發(fā)明的核酸構(gòu)建體包括抗原肽或所述抗原肽的片段的至少1、例如2、例如3、例如4、例如5、例如6、例如7、例如8、例如9、例如10、例如11、例如12、例如13、例如14、例如15、例如16、例如17、例如18、例如19、例如20種剪接變體。
超過一種抗原肽剪接變體可以包含等同或不等同的抗原肽。
在另一個(gè)實(shí)施方案中,根據(jù)本發(fā)明的核酸構(gòu)建體包括恒定鏈或其變體的至少1、例如2、例如3、例如4、例如5、例如6、例如7、例如8、例如9、例如10、例如11、例如12、例如13、例如14、例如15、例如16、例如17、例如18、例如19、例如20種剪接變體。
超過一種恒定鏈剪接變體可以包含等同或不等同恒定鏈或其變體。
在一個(gè)實(shí)施方案中,恒定鏈的至少一種剪接變體包括天然全長恒定鏈。在另一個(gè)實(shí)施方案中,恒定鏈的至少一種剪接變體包括恒定鏈的變體。在另外一個(gè)實(shí)施方案中,恒定鏈的至少一種剪接變體包括恒定鏈的變體,其中所述ii不包括ii-key區(qū)域的lrmk氨基酸殘基。在另一個(gè)實(shí)施方案中,恒定鏈的至少一種剪接變體包括恒定鏈的變體,其中所述ii不包括clip結(jié)構(gòu)域的m91和m99殘基。
由此可見剪接變體可以包括等同或不等同抗原肽和/或等同或不等同恒定鏈或其變體的任何組合。
以這種方式,可能的是“改組”(“shuffle”)包括恒定鏈的不同結(jié)構(gòu)域或區(qū)域的序列(外顯子),以便通過可變剪接獲得恒定鏈的變體。以這種方式,還可能的是“改組”包括一種或多種抗原肽的不同結(jié)構(gòu)域或區(qū)域的序列(外顯子),以便通過可變剪接獲得一種或多種所述抗原肽的變體。
恒定鏈
恒定鏈(ii)或mhcii類結(jié)合的恒定鏈或cd74或p31是非多態(tài)ii型嵌膜蛋白質(zhì),關(guān)于人和小鼠ii的氨基酸序列分別參見seqidnos:2和4,并且同樣地關(guān)于人和小鼠ii的核酸序列分別參見seqidnos:1和3。恒定鏈在淋巴細(xì)胞成熟和適應(yīng)性免疫應(yīng)答中具有多重功能,特別地靶向溶酶體區(qū)室是iiclip序列可以占據(jù)mhcii類分子,直至這些與胞內(nèi)體區(qū)室融合(pietersj.1997,curr.opin.immunol.,9:8996)。此外,ii已顯示發(fā)揮mhci類伴侶蛋白的功能(morris等人,2004,immunol.res.30:171-179),并且通過其胞內(nèi)體靶向序列,促進(jìn)針對(duì)共價(jià)連接抗原的cd4+而不是cd8+t細(xì)胞的增殖(diebold等人,2001,genether.8:487-493)。
恒定鏈蛋白質(zhì)包括幾個(gè)結(jié)構(gòu)域:包括信號(hào)或分選肽(也稱為溶酶體靶向序列)的細(xì)胞溶質(zhì)結(jié)構(gòu)域、跨膜結(jié)構(gòu)域、和其自身包括clip區(qū)域、key區(qū)域(包括lrmk殘基)的腔結(jié)構(gòu)域、核心結(jié)構(gòu)域和三聚結(jié)構(gòu)域。這些結(jié)構(gòu)域的兩側(cè)為高度彈性區(qū)域(strumptner-cuvelette&benaroch,2002,biochem.biophys.acta.,1542:1-13)。恒定鏈已在幾種生物中得到表征,包括脊椎動(dòng)物(例如雞)、哺乳動(dòng)物(例如牛、犬、小鼠和大鼠)和人。
本發(fā)明涉及包括這樣的序列的核酸構(gòu)建體,其中至少一種恒定鏈或其變體是生物特異性的,或可以與特定生物有關(guān)。優(yōu)選地,至少一種恒定鏈具有脊椎動(dòng)物起源,更優(yōu)選哺乳動(dòng)物起源且最優(yōu)選人起源。與之相關(guān),由seqidno:1限定的序列是來自人的恒定鏈的核酸序列。在另一個(gè)優(yōu)選實(shí)施方案中,至少一種恒定鏈具有禽起源,最優(yōu)選來自家雞(gallusgallusdomesticus)(雞)。在另外一個(gè)優(yōu)選實(shí)施方案中,至少一種恒定鏈衍生自魚,最優(yōu)選可以在養(yǎng)魚場中培育的魚(例如大麻哈魚或鮭魚)。在另外一個(gè)優(yōu)選實(shí)施方案中,至少一種恒定鏈衍生自雪貂。
采用的恒定鏈優(yōu)選是待接受核酸構(gòu)建體的生物的恒定鏈。本發(fā)明的一個(gè)目的是恒定鏈和宿主生物或處理接受者具有相同物種。
在本發(fā)明的一個(gè)實(shí)施方案中,包括至少一種恒定鏈或其變體的核酸構(gòu)建體具有下述前提:當(dāng)核酸構(gòu)建體包括至少一種恒定鏈的變體時(shí),所述恒定鏈不包括ii-key序列的lrmk氨基酸殘基。
在本發(fā)明的另一個(gè)實(shí)施方案中,包括至少一種恒定鏈或其變體的核酸構(gòu)建體具有下述前提:當(dāng)核酸構(gòu)建體包括至少一種恒定鏈的變體時(shí),所述恒定鏈包括ii-clip結(jié)構(gòu)域的變體。在一個(gè)實(shí)施方案中,所述變體是在位置91和99上的甲硫氨酸由另一個(gè)氨基酸的置換。在另一個(gè)實(shí)施方案中,所述變體是m91am99a雙點(diǎn)突變(在位置91和99上的氨基酸甲硫氨酸至丙氨酸的置換)。
在另一個(gè)實(shí)施方案中,ii變體是ii的前17個(gè)氨基酸的缺失(δ17ii)。
在第三個(gè)實(shí)施方案中,ii變體包括在位置91和99上的甲硫氨酸的置換,和ii的前17個(gè)氨基酸的缺失。
本發(fā)明人已驚訝地發(fā)現(xiàn)ii-key結(jié)構(gòu)域的lrmk殘基不是根據(jù)本發(fā)明用于引發(fā)免疫應(yīng)答所需的。
此外,本發(fā)明人已驚訝地發(fā)現(xiàn)在ii的位置91和99上的甲硫氨酸置換增加mhcii呈遞。
此外,本發(fā)明人已發(fā)現(xiàn)缺失ii的前17個(gè)氨基酸令人驚訝地增加記憶應(yīng)答。
本發(fā)明人已進(jìn)一步發(fā)現(xiàn)包括殘基編號(hào)50–118的恒定鏈的中心部分是獲得ii的完全效應(yīng)所需的。這種ii變體缺乏三聚結(jié)構(gòu)域。因此,在一個(gè)實(shí)施方案中,核酸構(gòu)建體包括至少一種恒定鏈或其變體,其中所述恒定鏈包括與來自另一種蛋白質(zhì)的三聚結(jié)構(gòu)域偶聯(lián)的氨基酸殘基編號(hào)50–118。所述其他蛋白質(zhì)可以是例如細(xì)菌蛋白質(zhì)或腺病毒纖維蛋白質(zhì)。
本發(fā)明還涉及核酸構(gòu)建體,其中編碼的至少一種恒定鏈?zhǔn)蔷哂兄辽?0個(gè)氨基酸的seqidno:2中鑒定的序列片段且與seqidno:2的相同片段具有至少85%同一性。
片段是來自如seqidno:2中所示恒定鏈的任何部分的至少40個(gè)氨基酸的片段。這包括包含下述殘基的片段:殘基1-40、10-50、20-60、25-65、30-70、35-75、40-80、45-85、50-90、55-95、60-100、65-105、70-110、75-115、80-120、85-125、90-130、95-135、100-140、105-145、110-150、115-155、120-160、125-165、130-170、135-175、140-180、145-185、150-190、155-195、160-200,165-205、170-210和175-216。它還包括如上文列出中的任何擴(kuò)大至其任一側(cè)的高達(dá)5個(gè)殘基的片段。它進(jìn)一步包括至少50個(gè)殘基、至少60個(gè)殘基、至少70個(gè)殘基、至少80個(gè)殘基、至少90個(gè)殘基、至少100個(gè)殘基、至少110個(gè)殘基、至少120個(gè)殘基、至少130個(gè)殘基、至少140個(gè)殘基、至少150個(gè)殘基、至少160個(gè)殘基、至少170個(gè)殘基、至少180個(gè)殘基至少190個(gè)殘基、至少200個(gè)殘基和至少210個(gè)殘基的片段。
在本發(fā)明的范圍內(nèi)包括與seqidno:2具有至少85%序列同一性、例如至少90%序列同一性、例如至少91%序列同一性、例如至少92%序列同一性、例如至少93%序列同一性、例如至少94%序列同一性、例如至少95%序列同一性、例如至少96%序列同一性、例如至少97%序列同一性、例如至少98%序列同一性、例如99%序列同一性的上述片段中的任何。
氨基酸序列之間的同一性/同源性可以使用眾所周知的評(píng)分矩陣進(jìn)行計(jì)算,例如blosum30、blosum40、blosum45、blosum50、blosum55、blosum60、blosum62、blosum65、blosum70、blosum75、blosum80、blosum85和blosum90中的任何一個(gè)。
在一個(gè)實(shí)施方案中,本發(fā)明是核酸構(gòu)建體,其中編碼的至少一種恒定鏈?zhǔn)蔷哂兄辽?86個(gè)氨基酸的seqidno:2的片段。這包括如上定義并且因此與seqidno:2的恒定鏈序列共享同一性的片段中的任何。
本發(fā)明另外涉及核酸構(gòu)建體,其中編碼的至少一種恒定鏈與seqidno:2至少85%等同。
這包含與seqidno:2具有至少85%序列同一性、例如至少90%序列同一性、例如至少91%序列同一性、例如至少92%序列同一性、例如至少93%序列同一性、例如至少94%序列同一性、例如至少95%序列同一性、例如至少96%序列同一性、例如至少97%序列同一性、例如至少98%序列同一性、例如99%序列同源性的衍生自如seqidno:2中提及的恒定鏈的任何序列,其包括在本發(fā)明的范圍內(nèi)。這包括比seqidno:2中所述的序列更長或更短的序列。
與起源、序列同一性或長度無關(guān),上述序列中的任何來自在此命名的恒定鏈變體。
由此可見在本發(fā)明的范圍內(nèi),來自任何生物的恒定鏈變體可以是根據(jù)上文的變體,即變體可以在ii-key區(qū)域中是改變的和/或在ii-clip區(qū)域中是改變的和/或是生物恒定鏈的片段和/或在恒定鏈的所有序列上或在其片段內(nèi)與所述恒定鏈至少85%等同。恒定鏈還可以來自生物的相關(guān)物種或來自遙遠(yuǎn)相關(guān)的物種。
本發(fā)明的另一個(gè)方面涉及如由核酸構(gòu)建體編碼的至少一種恒定鏈的區(qū)域、肽或結(jié)構(gòu)域的添加、去除或置換。這些區(qū)域、肽或結(jié)構(gòu)域中的一個(gè)或多個(gè)的去除將截短所得到的恒定鏈。區(qū)域、肽或結(jié)構(gòu)域的添加或替換包括從已知來源例如天然存在的蛋白質(zhì)或多肽或從人工合成的多肽或編碼其的核酸殘基中選擇這些序列的選項(xiàng)。區(qū)域、結(jié)構(gòu)域或肽的添加包括添加每種類型或不同類型區(qū)域、結(jié)構(gòu)域、肽中的1個(gè)、2個(gè)或更多個(gè),和編碼這些區(qū)域、結(jié)構(gòu)域和肽的核酸中的1個(gè)、2個(gè)、3個(gè)或更多個(gè)的選項(xiàng)。這些基于序列可以是彼此等同或不同的。區(qū)域、肽和結(jié)構(gòu)域無需與支架恒定鏈源于相同的生物。
區(qū)域、結(jié)構(gòu)域或肽的去除包括去除每種類型或不同類型區(qū)域、結(jié)構(gòu)域、肽中的1個(gè)、2個(gè)、3個(gè)或更多個(gè)和去除編碼這些區(qū)域、結(jié)構(gòu)域和肽的氨基酸殘基中的1個(gè)、2個(gè)、3個(gè)、4個(gè)、5個(gè)、6個(gè)、7個(gè)、8個(gè)、9個(gè)、10個(gè)、11個(gè)、12個(gè)、13個(gè)、14個(gè)、15個(gè)、16個(gè)、17個(gè)、18個(gè)、19個(gè)、20個(gè)或更多個(gè)的選項(xiàng)。執(zhí)行編碼所得到的多肽的個(gè)別核苷酸以及核苷酸段的添加、缺失和置換是本領(lǐng)域眾所周知的。
當(dāng)確定哪些置換、缺失、重排或其他改變對(duì)于構(gòu)建體將是有利的時(shí),比對(duì)來自不同生物的同源基因或蛋白質(zhì)的核酸且特別是蛋白質(zhì)序列可以具有極大幫助。如下舉例說明的比對(duì)人和鼠類恒定鏈序列給出哪些氨基酸殘基對(duì)于在這些生物中恒定鏈的結(jié)構(gòu)和功能可以具有重要性的指示–這些是在2個(gè)序列之間是保守的殘基。同樣地,推測的較不重要的殘基是在其中序列不同的那些。就本發(fā)明而言有利的是執(zhí)行變體殘基/區(qū)域的置換和/或缺失。當(dāng)嘗試突變或缺失或以其他方式改變例如人恒定鏈的序列以便改善其免疫應(yīng)答刺激能力時(shí),檢查保守殘基且制備例如同源置換(即其中氨基酸被視為具有例如相同結(jié)構(gòu)質(zhì)量、極性、疏水性或其他的置換)也可以是相關(guān)的。
在衍生自人和小鼠的恒定鏈蛋白質(zhì)如下比對(duì)中,key區(qū)域的lrmk氨基酸殘基是具有下劃線的。
key區(qū)域
本發(fā)明的一個(gè)優(yōu)選實(shí)施方案涉及至少一種恒定鏈的key區(qū)域的去除、置換或替換。如上所述,key區(qū)域的添加或替換包括用來自相同或其他生物的恒定鏈的key區(qū)域或來自相同或其他生物的key區(qū)域的變體來添加或替換在選擇的一種或多種恒定鏈的變體中的現(xiàn)有key區(qū)域的選項(xiàng)。如由上文可見的,變體key區(qū)域可以是特異性生成的key區(qū)域的突變體形式,通過單個(gè)或多個(gè)核酸置換、缺失或添加生成。一個(gè)優(yōu)選實(shí)施方案包括單獨(dú)的key區(qū)域的n末端或c末端相鄰序列,但不含key區(qū)域自身。相鄰意指在key區(qū)域的10個(gè)殘基內(nèi)、在key區(qū)域的20個(gè)殘基內(nèi)、在30個(gè)殘基內(nèi)、在40個(gè)殘基內(nèi)、在50個(gè)殘基內(nèi)、在75個(gè)殘基內(nèi)或在100個(gè)殘基內(nèi)的任何氨基酸。
一個(gè)最優(yōu)選的實(shí)施方案包括key區(qū)域的一個(gè)或多個(gè)置換或缺失,導(dǎo)致在key區(qū)域中包括的lrmk氨基酸中的1個(gè)、2個(gè)、3個(gè)或4個(gè)氨基酸殘基的置換或缺失。在一個(gè)實(shí)施方案中,缺失在key區(qū)域中包括的lrmk氨基酸中的至少1個(gè)、例如2個(gè)、例如3個(gè)、例如4個(gè)。在另一個(gè)實(shí)施方案中,在key區(qū)域中包括的lrmk氨基酸中的至少1個(gè)、例如2個(gè)、例如3個(gè)、例如4個(gè)由其他氨基酸置換。所述氨基酸可以是選自下述的任何氨基酸:g(甘氨酸)、p(脯氨酸)、a(丙氨酸)、v(纈氨酸)、l(亮氨酸)、i(異亮氨酸)、m(甲硫氨酸)、c(半胱氨酸)、f(苯丙氨酸)、y(酪氨酸)、w(色氨酸)、h(組氨酸)、k(賴氨酸)、r(精氨酸)、q(谷氨酰胺)、n(天冬酰胺)、e(谷氨酸)、d(天冬氨酸)、s(絲氨酸)和t(蘇氨酸)。
在一個(gè)特定實(shí)施方案中,lrmk氨基酸殘基各自由丙氨酸(a)氨基酸殘基置換,從而序列閱讀為:aaaa。在另一個(gè)實(shí)施方案中,lrmk氨基酸殘基由包括同義或等值置換的氨基酸置換。例如,氨基酸殘基l可以由i、v、m或f置換;r可以由k、h、e或d置換;m可以由l、i、f或v置換;并且k可以由h或r置換。
在本發(fā)明的一個(gè)實(shí)施方案中,key區(qū)域可以包括大于lrmk殘基,或lrmk殘基可以由大于4個(gè)氨基酸殘基的序列替換。
本發(fā)明的一個(gè)實(shí)施方案涉及如上所述不含key區(qū)域的恒定鏈的片段。這些片段可以是至少5個(gè)氨基酸殘基長,長度至少10個(gè)殘基、至少15個(gè)殘基、至少20個(gè)殘基、至少25個(gè)殘基、至少30個(gè)殘基或至少35個(gè)殘基。另一個(gè)實(shí)施方案涉及恒定鏈的片段,其中信號(hào)肽被去除,并且恒定鏈片段是至少10個(gè)氨基酸殘基長,長度至少15個(gè)殘基、至少20個(gè)殘基、至少25個(gè)殘基、至少30個(gè)殘基、至少35個(gè)殘基、至少50個(gè)殘基、至少60個(gè)殘基、至少70個(gè)殘基、至少80個(gè)殘基、至少90個(gè)殘基、至少100個(gè)殘基、至少110個(gè)殘基、至少120個(gè)殘基、至少130個(gè)殘基、至少140個(gè)殘基、至少150個(gè)殘基、至少160個(gè)殘基、至少170個(gè)殘基或至少180個(gè)殘基。
在本發(fā)明的一個(gè)實(shí)施方案中,由如本文描述的核酸構(gòu)建體編碼的至少一種恒定鏈不包括ii-key區(qū)域的lrmk氨基酸殘基。
在一個(gè)實(shí)施方案中,本發(fā)明因此涉及包括與至少一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段連接的至少一種恒定鏈或其變體的核酸構(gòu)建體,其中所述恒定鏈或其變體不包括ii-key區(qū)域的lrmk氨基酸殘基。
clip區(qū)域
本發(fā)明的另一個(gè)實(shí)施方案涉及至少一種恒定鏈的clip區(qū)域的去除、添加或替換。如上所述,clip區(qū)域的添加或替換包括用來自相同或其他生物的恒定鏈的clip區(qū)域或來自相同或其他生物的clip區(qū)域的變體來添加或替換在選擇的一種或多種恒定鏈的變體中的現(xiàn)有clip區(qū)域的選項(xiàng)。如由上文可見的,變體clip區(qū)域可以是特異性生成的clip區(qū)域的突變體形式,通過單個(gè)或多個(gè)核酸置換、缺失或添加生成。一個(gè)優(yōu)選實(shí)施方案包括單獨(dú)的clip區(qū)域,或連同n末端相鄰序列或c末端相鄰序列的clip區(qū)域,不含恒定鏈的任何其他區(qū)域或結(jié)構(gòu)域。其他優(yōu)選實(shí)施方案包括單獨(dú)的clip區(qū)域的n末端或c末端相鄰序列,但不含clip區(qū)域自身。相鄰意指在clip區(qū)域的10個(gè)殘基內(nèi)、在clip區(qū)域的20個(gè)殘基內(nèi)、在30個(gè)殘基內(nèi)、在40個(gè)殘基內(nèi)、在50個(gè)殘基內(nèi)、在75個(gè)殘基內(nèi)或在100個(gè)殘基內(nèi)的任何氨基酸,。
一個(gè)優(yōu)選實(shí)施方案包括clip區(qū)域的一個(gè)或多個(gè)置換或缺失,導(dǎo)致在clip區(qū)域的1個(gè)、2個(gè)、3個(gè)、4個(gè)或更多個(gè)氨基酸殘基的置換或缺失。在一個(gè)實(shí)施方案中,缺失在clip區(qū)域中包括的氨基酸中的至少1個(gè)、例如2個(gè)、例如3個(gè)、例如4個(gè)或更多個(gè)。在另一個(gè)實(shí)施方案中,在clip區(qū)域中包括的氨基酸中的至少1個(gè)、例如2個(gè)、例如3個(gè)、例如4個(gè)或更多個(gè)由其他氨基酸置換。所述氨基酸可以是選自下述的任何氨基酸:g(甘氨酸)、p(脯氨酸)、a(丙氨酸)、v(纈氨酸)、l(亮氨酸)、i(異亮氨酸)、m(甲硫氨酸)、c(半胱氨酸)、f(苯丙氨酸)、y(酪氨酸)、w(色氨酸)、h(組氨酸)、k(賴氨酸)、r(精氨酸)、q(谷氨酰胺)、n(天冬酰胺)、e(谷氨酸)、d(天冬氨酸)、s(絲氨酸)和t(蘇氨酸)。
本發(fā)明的一個(gè)實(shí)施方案涉及如上所述不含clip區(qū)域的恒定鏈的片段。這些片段可以是至少5個(gè)氨基酸殘基長,長度至少10個(gè)殘基、至少15個(gè)殘基、至少20個(gè)殘基、至少25個(gè)殘基、至少30個(gè)殘基或至少35個(gè)殘基。另一個(gè)實(shí)施方案涉及恒定鏈的片段,其中信號(hào)肽被去除,并且恒定鏈片段是至少10個(gè)氨基酸殘基長,長度至少15個(gè)殘基、至少20個(gè)殘基、至少25個(gè)殘基、至少30個(gè)殘基、至少35個(gè)殘基、至少50個(gè)殘基、至少60個(gè)殘基、至少70個(gè)殘基、至少80個(gè)殘基、至少90個(gè)殘基、至少100個(gè)殘基、至少110個(gè)殘基、至少120個(gè)殘基、至少130個(gè)殘基、至少140個(gè)殘基、至少150個(gè)殘基、至少160個(gè)殘基、至少170個(gè)殘基或至少180個(gè)殘基。
在一個(gè)特定實(shí)施方案中,在位置91和99上的m氨基酸殘基各自由丙氨酸(a)氨基酸殘基置換,從而序列閱讀為:m91am99a。在另一個(gè)實(shí)施方案中,在位置91和99上的m氨基酸殘基由包括同義或等值置換的氨基酸置換。
在本發(fā)明的一個(gè)實(shí)施方案中,由如本文描述的核酸構(gòu)建體編碼的至少一種恒定鏈不包括在ii-clip序列的位置91和99上的m氨基酸殘基。
在一個(gè)實(shí)施方案中,本發(fā)明因此涉及包括與至少一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段連接的至少一種恒定鏈或其變體的核酸構(gòu)建體,其中所述恒定鏈或其變體不包括在ii-clip區(qū)域的位置91和99上的m氨基酸殘基。
n或c末端改變:
本發(fā)明的一個(gè)實(shí)施方案涉及至少一種恒定鏈的n或c末端區(qū)域的去除(缺失)、置換或替換。如上所述,n或c末端區(qū)域的添加或替換包括用來自相同或其他生物的恒定鏈或其他蛋白質(zhì)的n或c末端區(qū)域或來自相同或其他生物的n或c末端區(qū)域的變體來添加或替換在選擇的一種或多種恒定鏈的變體中的現(xiàn)有n或c末端區(qū)域的選項(xiàng)。如由上文可見的,變體n或c末端區(qū)域可以是特異性生成的n或c末端區(qū)域的突變體形式,其通過單個(gè)或多個(gè)核酸置換、缺失或添加生成。
一個(gè)實(shí)施方案包括ii的前(n末端)或末(c末端)氨基酸的缺失,例如ii的前或末1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、125、130、135、140、145或150個(gè)氨基酸。
在本發(fā)明的一個(gè)特定實(shí)施方案中,ii變體包括ii的前17個(gè)氨基酸的缺失(δ17ii)。
本發(fā)明的一個(gè)實(shí)施方案涉及不含完整n或c末端區(qū)域的如上所述恒定鏈的片段。這些片段可以是至少5個(gè)氨基酸殘基長,長度至少10個(gè)殘基、至少15個(gè)殘基、至少20個(gè)殘基、至少25個(gè)殘基、至少30個(gè)殘基或至少35個(gè)殘基。另一個(gè)實(shí)施方案涉及恒定鏈的片段,其中信號(hào)肽被去除,并且恒定鏈片段是至少10個(gè)氨基酸殘基長,長度至少15個(gè)殘基、至少20個(gè)殘基、至少25個(gè)殘基、至少30個(gè)殘基、至少35個(gè)殘基、至少50個(gè)殘基、至少60個(gè)殘基、至少70個(gè)殘基、至少80個(gè)殘基、至少90個(gè)殘基、至少100個(gè)殘基、至少110個(gè)殘基、至少120個(gè)殘基、至少130個(gè)殘基、至少140個(gè)殘基、至少150個(gè)殘基、至少160個(gè)殘基、至少170個(gè)殘基或至少180個(gè)殘基。
在本發(fā)明的一個(gè)實(shí)施方案中,ii的跨膜區(qū)段的所有或部分可以由來自任何其他蛋白質(zhì)例如趨化因子受體ccr6tm6的相應(yīng)區(qū)段替換。
在本發(fā)明的另一個(gè)實(shí)施方案中,ii的跨膜區(qū)段的所有或部分可以由來自趨化因子受體ccr6tm6的相應(yīng)區(qū)段替換。
信號(hào)肽
本發(fā)明的另一個(gè)實(shí)施方案涉及至少一種恒定鏈,其中信號(hào)肽被去除、替換或添加到編碼恒定鏈的序列上。信號(hào)肽是決定蛋白質(zhì)在細(xì)胞中的最終定位的短氨基酸序列,也被稱為分選肽。決定蛋白質(zhì)至亞細(xì)胞區(qū)室例如內(nèi)質(zhì)網(wǎng)、高爾基體和各種區(qū)室包括高爾基體、核、質(zhì)膜、線粒體及其中的各種空間和膜、過氧化物酶體、溶酶體、胞內(nèi)體和分泌囊泡的信號(hào)肽都包括在本發(fā)明的范圍內(nèi)。一個(gè)優(yōu)選實(shí)施方案包括單獨(dú)的恒定鏈的溶酶體靶向序列。
抗原
恒定鏈的上述變體中的任何以這樣的形式包含在本發(fā)明中,其中所述變體中的至少一種與至少一種抗原例如抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段可操作地連接。
本發(fā)明的一個(gè)目的是包括但不限于源于致病性生物的抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的片段、癌癥特異性多肽和抗原、和與異常生理學(xué)應(yīng)答相關(guān)的蛋白質(zhì)或肽。
更優(yōu)選地,本發(fā)明的一個(gè)目的是包括源于下述類型病原體中的任何的抗原:病毒、微生物和寄生蟲。這包括任何動(dòng)物已知的病原體。優(yōu)選具有來自哺乳動(dòng)物病原體的抗原,例如特異性靶向哺乳動(dòng)物例如雪貂的病原體。優(yōu)選具有來自人病原體的抗原。一般而言,可以使用發(fā)現(xiàn)與人病原體或疾病相關(guān)的任何抗原。
在另一個(gè)實(shí)施方案中,優(yōu)選具有來自禽病原體的抗原,例如特異性靶向鳥或家禽的病原體。更優(yōu)選具有來自雞(家雞)的抗原。一般而言,可以使用發(fā)現(xiàn)與禽病原體相關(guān)的任何抗原。
在另外一個(gè)實(shí)施方案中,優(yōu)選具有來自魚類病原體即特異性靶向魚的病原體的抗原。更優(yōu)選具有來自可以在養(yǎng)魚場中培育的魚的抗原。一般而言,可以使用發(fā)現(xiàn)與魚類病原體相關(guān)的任何抗原。
病毒抗原
在一個(gè)優(yōu)選實(shí)施方案中,至少一種抗原可以源于但不限于下述病毒科中的任何:腺病毒、沙粒病毒科、星狀病毒科、布尼亞病毒科、嵌杯病毒科、冠狀病毒科、黃病毒科、皰疹病毒科、正粘病毒科、副粘病毒科、細(xì)小rna病毒科、痘病毒科、呼腸病毒科、逆轉(zhuǎn)錄病毒科、彈狀病毒科和披膜病毒科。
更具體而言,至少一種抗原或抗原序列可以衍生自下述病毒中的任何:甲型流感例如h1n1、h1n2、h3n2和h5n1(禽流感),乙型流感,丙型流感病毒,甲型肝炎病毒,乙型肝炎病毒,丙型肝炎病毒,丁型肝炎病毒、戊型肝炎病毒,輪狀病毒屬,諾瓦克病毒屬中的任何病毒,腸腺病毒,細(xì)小病毒屬,登革熱病毒,猴痘,單股負(fù)鏈病毒目,狂犬病病毒屬例如狂犬病病毒,拉戈斯蝙蝠病毒,莫科拉病毒,杜溫哈格病毒,歐洲蝙蝠病毒1和2以及澳大利亞蝙蝠病毒,短暫熱病毒屬,水泡病毒屬,水皰性口炎病毒(vsv),皰疹病毒例如單純皰疹病毒1和2型,水泡帶狀皰疹,巨細(xì)胞病毒,eb病毒(ebv),人皰疹病毒(hhv),人皰疹病毒6和8型,人免疫缺陷病毒(hiv),乳頭狀瘤病毒,鼠類γ皰疹病毒,沙粒病毒例如阿根廷出血熱病毒、玻利維亞出血熱病毒、sabia伴隨出血熱病毒、委內(nèi)瑞拉出血熱病毒、拉沙熱病毒,machupo病毒,淋巴細(xì)胞性脈絡(luò)叢腦膜炎病毒(lcmv),布尼亞病毒科例如克里米亞-剛果出血熱病毒,漢他病毒,引起伴腎病綜合征的出血熱病毒,裂谷熱病毒,絲狀病毒科(絲狀病毒)包括埃博拉出血熱和馬爾堡出血熱,黃病毒科包括科薩努爾森林病病毒,鄂木斯克出血熱病毒,引起蜱傳腦炎的病毒和副粘病毒例如亨德拉病毒和尼帕病毒,重型天花和輕型天花(天花),甲病毒例如委內(nèi)瑞拉馬腦炎病毒,東方馬腦炎病毒,西方馬腦炎病毒,sars-相關(guān)冠狀病毒(sars-cov),西尼羅河病毒,引起任何腦炎的病毒。
在本發(fā)明的一個(gè)優(yōu)選實(shí)施方案中,至少一種抗原蛋白質(zhì)或肽來自選自下述的病毒:hiv、丙型肝炎病毒、流感病毒、皰疹病毒、拉沙、埃博拉、天花、禽流感、絲狀病毒、馬爾堡和乳頭狀瘤病毒。
在本發(fā)明的一個(gè)更優(yōu)選實(shí)施方案中,至少一種抗原蛋白質(zhì)或肽選自下述和/或可以是下述中的任何的至少一個(gè)抗原片段:水皰性口炎病毒糖蛋白(vsv-gp);甲型流感ns-1(非結(jié)構(gòu)蛋白質(zhì)1)、m1(基質(zhì)蛋白質(zhì)1)、np(核蛋白)、nep、m2、m2e、ha、na、pa、pb1、pb2、pb1-f2;lcmvnp、lcmvgp;埃博拉gp、埃博拉np;hiv抗原tat、vif、rev、vpr、gag、pol、nef、env、vpu;siv抗原tat、vif、rev、vpr、gag、pol、nef、env;鼠類γ皰疹病毒m2、m3和orf73(例如mhv-68m2、m3和orf73);雞卵白蛋白(ova);或輔助t細(xì)胞表位。將本文提及抗原中的任何的2種或更多種組合是在本發(fā)明的范圍內(nèi)的。
微生物抗原
本發(fā)明的一個(gè)實(shí)施方案包括來自微生物的至少一種抗原蛋白質(zhì)或肽或抗原蛋白質(zhì)或肽的片段。更具體而言,至少一種抗原可以衍生自來自非詳盡列表的下述中的一種:炭疽(炭疽芽孢桿菌(bacillusanthracis)),結(jié)核分枝桿菌(mycobacteriumtuberculosis),沙門氏菌屬(salmonella)(雞沙門氏菌(salmonellagallinarum)、雞白痢沙門氏菌(s.pullorum)、傷寒沙門氏菌(s.typhi)、腸炎沙門氏菌(s.enteridtidis)、副傷寒沙門氏菌(s.paratyphi)、都柏林沙門氏菌(s.dublin)、鼠傷寒沙門氏菌(s.typhimurium)),肉毒梭菌(clostridiumbotulinum),產(chǎn)氣莢膜梭菌(clostridiumperfringens),白喉?xiàng)U菌(corynebacteriumdiphtheriae),百日咳博德特氏菌(bordetellapertussis),彎曲桿菌屬(campylobacter)例如空腸彎曲桿菌(campylobacterjejuni),新型隱球菌(crytococcusneoformans),鼠疫耶爾森氏菌(yersiniapestis),小腸結(jié)腸炎耶爾森氏菌(yersiniaenterocolitica),假結(jié)核耶爾森氏菌(yersiniapseudotuberculosis),單核細(xì)胞增多性李斯特菌(listeriamonocytogenes),鉤端螺旋體屬(leptospira)物種,嗜肺軍團(tuán)菌(legionellapneumophila),伯氏疏螺旋體(borreliaburgdorferi),鏈球菌屬(streptococcus)物種例如肺炎鏈球菌(streptococcuspneumoniae),腦膜炎奈瑟球菌(neisseriameningitides),流感嗜血桿菌(haemophilusinfluenzae),弧菌屬(vibrio)物種例如霍亂弧菌(vibriocholerae)o1、霍亂弧菌非o1、副溶血弧菌(v.parahaemolyticus)、副溶血弧菌、溶藻弧菌(v.alginolyticus)、弗尼斯弧菌(v.furnissii)、鯊魚弧菌(v.carchariae)、霍氏弧菌(v.hollisae)、辛辛那提弧菌(v.cincinnatiensis)、梅氏弧菌(v.metschnikovii)、海魚弧菌(v.damsela)、擬態(tài)弧菌(v.mimicus)、河流弧菌(v.fluvialis)、創(chuàng)傷弧菌(v.vulnificus),蠟狀芽孢桿菌(bacilluscereus),嗜水氣單胞菌(aeromonashydrophila)、豚鼠氣單胞菌(aeromonascaviae)、溫和氣單胞菌(aeromonassobria)和維氏氣單胞菌(aeromonasveronii),類志賀鄰單胞菌(plesiomonasshigelloides),志賀氏菌屬(shigella)物種例如宋氏志賀氏菌(shigellasonnei)、鮑氏志賀氏菌(s.boydii)、弗氏志賀氏菌(s.flexneri)和痢疾志賀氏菌(s.dysenteriae),致瀉大腸埃希氏菌(enterovirulentescherichiacoli)eec(產(chǎn)腸毒素大腸埃希氏菌(escherichiacoli-enterotoxigenic)(etec)、腸致病性大腸埃希氏菌(escherichiacoli-enteropathogenic)(epec)、腸出血性大腸埃希氏菌o157∶h7(escherichiacolio157:h7enterohemorrhagic)(ehec)、腸侵襲性大腸埃希氏菌(escherichiacoli-enteroinvasive)(eiec)),葡萄球菌屬(staphylococcus)物種例如金黃色葡萄球菌(s.aureus)且特別是萬古霉素中間產(chǎn)物/抗性物種(visa/vrsa)或多藥抗性物種(mrsa),志賀氏菌屬物種例如弗氏志賀氏菌、宋氏志賀氏菌、痢疾志賀氏菌,微小隱孢子蟲(cryptosporidiumparvum),布魯氏菌屬(brucella)物種例如流產(chǎn)布魯氏菌(b.abortus)、馬爾他布魯氏菌(b.melitensis)、綿羊布魯氏菌(b.ovis)、豬布魯氏菌(b.suis)和犬布魯氏菌(b.canis),鼻疽伯克霍爾德菌(burkholderiamallei)和類鼻疽伯克霍爾德菌(burkholderiapseudomallei),鸚鵡衣原體(chlamydiapsittaci),貝氏柯克斯體(coxiellaburnetii),土拉熱弗朗西絲菌(francisellatularensis),普氏立克次體(rickettsiaprowazekii),夾膜組織胞漿菌(histoplasmacapsulatum),粗球孢子菌(coccidioidesimmitis)。
在本發(fā)明的一個(gè)優(yōu)選實(shí)施方案中,至少一種抗原蛋白質(zhì)或肽來自選自下述的微生物:結(jié)核分枝桿菌、炭疽芽孢桿菌、葡萄球菌屬物種和弧菌屬物種。
寄生蟲抗原
本發(fā)明的一個(gè)實(shí)施方案涉及核酸構(gòu)建體,其中編碼的至少一種抗原蛋白質(zhì)或肽來自寄生蟲。
本發(fā)明的另一個(gè)實(shí)施方案涉及核酸構(gòu)建體,其包括來自上述病原體中的任何的至少2種抗原蛋白質(zhì)或肽的組合。
優(yōu)選地,抗原衍生自但不限于選自下述的寄生蟲:瘧原蟲屬(plasmodium)物種例如三日瘧原蟲(plasmodiummalariae)、卵形瘧原蟲(plasmodiumovale)、間日瘧原蟲(plasmodiumvivax)、惡性瘧原蟲(plasmodiumfalciparum),微小內(nèi)蜒阿米巴(endolimaxnana),藍(lán)氏賈第蟲(giardialamblia),溶組織內(nèi)阿米巴(entamoebahistolytica),微小隱孢子蟲(cryptosporidiumparvum),人芽囊原蟲(blastocystishominis),陰道毛滴蟲(trichomonasvaginalis),剛地弓形蟲(toxoplasmagondii),圓孢子球蟲(cyclosporacayetanensis),鼠隱孢子蟲(cryptosporidiummuris),卡氏肺囊蟲(pneumocystiscarinii),杜氏利什曼原蟲(leishmaniadonovani)、熱帶利什曼原蟲(leishmaniatropica)、巴西利什曼原蟲(leishmaniabraziliensis)、墨西哥利什曼原蟲(leishmaniamexicana),棘阿米巴屬(acanthamoeba)物種例如卡氏棘阿米巴(acanthamoebacastellanii)和卡伯特森氏棘阿米巴(a.culbertsoni),福氏耐格里原蟲(naegleriafowleri),克魯斯氏錐蟲(trypanosomacruzi)、布氏羅得西亞錐蟲(trypanosomabruceirhodesiense)、布氏岡比亞錐蟲(trypanosomabruceigambiense),貝氏等孢子球蟲(isosporabelli),結(jié)腸小袋纖毛蟲(balantidiumcoli),蛔蟲(roundworm)(似蚓蛔線蟲(ascarislumbricoides)),鉤蟲(hookworm)(美洲鉤蟲(necatoramericanus)、十二指腸鉤蟲(ancylostomaduodenal)),蟯蟲(pinworm)(蠕形住腸線蟲(enterobiusvermicularis)),蛔蟲(犬弓蛔蟲(toxocaracanis)、貓弓蛔蟲(toxocaracati)),犬心絲蟲(heartworm)(犬惡絲蟲(dirofilariaimmitis)),類圓線蟲屬(strongyloides)(糞類圓線蟲(stronglyoidesstercoralis)),毛線蟲屬(trichinella)(旋毛線蟲(trichinellaspiralis)),絲蟲屬(filaria)(班氏吳策線蟲(wuchereriabancrofti)、馬來絲蟲(brugiamalayi)、旋盤尾絲蟲(onchocercavolvulus)、羅阿絲蟲(loaloa)、鏈尾曼森線蟲(mansonellastreptocerca)、常現(xiàn)曼森線蟲(mansonellaperstans)、奧氏曼森線蟲(mansonellaozzardi))和異尖線蟲(anisakine)幼蟲(簡單異尖線蟲(anisakissimplex)(鯡魚蠕蟲)),擬地新線蟲屬(pseudoterranova)(海豹線蟲(phocanema)、鉆線蟲(terranova))形態(tài)種(鱈魚或海豹蠕蟲),對(duì)盲囊線蟲屬(contracaecum)物種和宮脂線蟲(hysterothylacium)(鮪蛔線蟲屬(thynnascaris)物種),鞭形鞭蟲(trichuristrichiura),牛肉絳蟲(beeftapeworm)(牛肉絳蟲(taeniasaginata)),豬肉絳蟲(porktapeworm)(豬帶絳蟲(taeniasolium)),魚絳蟲(fishtapeworm)(闊節(jié)裂頭絳蟲(diphyllobothriumlatum))和犬絳蟲(dogtapeworm)(犬復(fù)孔絳蟲(dipylidiumcaninum)),腸吸蟲(intestinalfluke)(布氏姜片吸蟲(fasciolopsisbuski)),血吸蟲(bloodfluke)(日本血吸蟲(schistosomajaponicum)、曼氏血吸蟲(schistosomamansoni)、埃及血吸蟲(schistosomahaematobium)),肝吸蟲(liverfluke)(華支睪吸蟲(clonorchissinensis)),東方肺吸蟲(orientallungfluke)(衛(wèi)氏并殖吸蟲(paragonimuswestermani))和綿羊肝吸蟲(sheepliverfluke)(肝片形吸蟲(fasciolahepatica)),鮭隱孔吸蟲(nanophyetussalmincola)和希氏侏形吸蟲(n.schikhobalowi)。
在本發(fā)明的一個(gè)優(yōu)選實(shí)施方案中,至少一種抗原蛋白質(zhì)或肽來自選自下述的寄生蟲:瘧原蟲屬物種、利什曼原蟲屬物種和錐蟲屬物種。
本發(fā)明的至少一種抗原可以是來自惡性瘧原蟲的var2csa。在本發(fā)明的一個(gè)優(yōu)選實(shí)施方案中,至少一種抗原蛋白質(zhì)或肽或抗原蛋白質(zhì)或肽的片段是var2csa。
家畜抗原
本發(fā)明的一個(gè)方面涉及衍生自感染家畜的疾病或因子的抗原和/或抗原序列,特別是商業(yè)上相關(guān)的動(dòng)物例如豬、牛、馬、綿羊、山羊、美洲駝、兔、貂、小鼠、大鼠、犬、貓、雪貂,家禽例如雞、火雞、雉雞及其他,魚例如鮭魚、大麻哈魚、鱈魚及其他養(yǎng)殖物種。至少一種抗原或抗原序列可以衍生自其的疾病或其因子(agent)的例子包括但不限于:多物種疾病例如:炭疽、aujeszky氏病、藍(lán)舌病,布魯氏菌病例如:流產(chǎn)布魯氏菌、馬爾他布魯氏菌或豬布魯氏菌;克里米亞剛果出血熱,棘球蚴病/包蟲病,細(xì)小rna病毒科的病毒,引起口蹄疫的口蹄疫病毒屬特別是7個(gè)免疫上不同的血清型中的任何:a、o、c、sat1、sat2、sat3、asia1,或心水病,日本腦炎,鉤端螺旋體病,新大陸螺旋蠅蛆病(嗜人錐蠅(cochliomyiahominivorax)),舊大陸螺旋蠅蛆病(蛆癥金蠅(chrysomyabezziana)),副結(jié)核,q型熱,狂犬病,裂谷熱,牛疫,旋毛蟲病,土拉菌病,水皰性口腔炎或西尼羅河熱;牛疾病例如:牛邊蟲病、牛巴貝蟲病、牛生殖道彎曲桿菌病、牛海綿狀腦病、牛結(jié)核病、牛病毒性腹瀉、牛傳染性胸膜肺炎、地方流行性牛白血病、出血性敗血癥、牛傳染性鼻氣管炎/傳染性膿皰性外陰陰道炎、疙瘩皮膚病(lumpkyskindisease)、惡性卡他熱、泰勒蟲病、毛滴蟲病或錐蟲病(舌蠅傳播的);綿羊和山羊疾病例如:山羊關(guān)節(jié)炎/腦炎、接觸傳染性無乳癥、接觸傳染性山羊胸膜肺炎、地方流行性母羊流產(chǎn)(綿羊衣原體病)、梅迪-維斯納病(maedi-visna)、內(nèi)羅畢綿羊病、綿羊附睪炎(羊布魯氏菌)、小反芻動(dòng)物瘟疫、沙門氏菌病(羊流產(chǎn)沙門氏菌(s.abortusovis))、羊瘙癢癥、綿羊痘和山羊痘;馬疾病例如:非洲馬瘟病、馬接觸傳染性子宮炎、馬交媾病、馬腦脊髓炎(東方)、馬腦脊髓炎(西方)、馬傳染性貧血、馬流感、馬焦蟲病、馬鼻肺炎、馬病毒性動(dòng)脈炎、馬鼻疽、蘇拉病(伊氏錐蟲(trypanosomaevansi))或委內(nèi)瑞拉馬腦脊髓炎;豬疾病例如:非洲豬瘟、古典豬瘟、尼帕病毒性腦炎、豬囊蟲病、豬繁殖與呼吸綜合征、豬水皰病或傳染性腸胃炎;禽疾病例如:禽衣原體病、禽傳染性支氣管炎、禽傳染性喉氣管炎、禽支原體病(雞毒支原體(m.gallisepticum))、禽支原體病(滑液支原體(m.synoviae))、鴨病毒性肝炎、家禽霍亂、家禽傷寒、高致病性禽流感這是任何甲型或乙型流感病毒且特別是h5n1、傳染性法氏囊病(gumboro病)、馬立克氏病、新城疫、雞白痢或火雞鼻氣管炎;兔類動(dòng)物和嚙齒類動(dòng)物疾病例如:病毒性腸炎、粘液瘤病或兔出血性疾病;魚疾病例如:地方流行性造血器官壞死癥、傳染性造血器官壞死癥、鯉魚春季病毒血癥、病毒性出血性敗血癥、傳染性胰臟壞死病、傳染性鮭魚貧血、流行性潰瘍綜合征、細(xì)菌性腎病(鮭魚腎桿菌(renibacteriumsalmoninarum))、三代蟲病(gyrodactylosis)(三代蟲病(gyrodactylussalaris))、紅海鯛虹彩病毒病;或其他疾病例如駱駝痘或利什曼病。
在本發(fā)明的一個(gè)優(yōu)選實(shí)施方案中,至少一種抗原蛋白質(zhì)或肽來自aujeszky氏病、口蹄疫、水皰性口炎病毒、禽流感或新城疫。
本發(fā)明的另外一個(gè)優(yōu)選實(shí)施方案涉及至少一種抗原蛋白質(zhì)或肽或所述抗原蛋白質(zhì)或肽的片段,其是與上述抗原中的任何具有至少85%同一性的抗原肽或蛋白質(zhì)。氨基酸之間的同源性或同一性可以通過先前提及的blosum評(píng)分矩陣中的任何進(jìn)行計(jì)算。
癌抗原
許多蛋白質(zhì)/糖蛋白已得到鑒定且與特定類型的癌癥聯(lián)系;這些被稱為癌癥特異性多肽、腫瘤相關(guān)抗原或癌抗原。一般而言,可以使用發(fā)現(xiàn)與癌癥腫瘤相關(guān)的任何抗原。其中可以發(fā)現(xiàn)癌癥特異性抗原的一種方法是通過扣除分析(subtractionanalysis)例如各種微陣列分析,例如dna微陣列分析。此處,比較在健康和癌性患者之間、在癌性患者組之間、或在相同患者中的健康和癌性組織之間的基因表達(dá)模式(如由所述基因編碼的rna或蛋白質(zhì)水平中可見的)。從彼此中“扣除”具有大約相等的表達(dá)水平的基因,留下在健康和癌性組織之間不同的基因/基因產(chǎn)物。這種方法是本領(lǐng)域已知的,并且可以用作鑒定新型癌抗原的方法或產(chǎn)生對(duì)于給定患者或患者組特異的基因表達(dá)概況。因此鑒定的抗原,單一抗原和它們可以在其中發(fā)現(xiàn)的組合落入本發(fā)明的范圍內(nèi)。
優(yōu)選地,本發(fā)明的至少一種抗原衍生自但不限于選自下述的癌癥特異性多肽:mage-3,mage-1,gp100,gp75,trp-2,酪氨酸酶,mart-1,cea,ras,p53,b-連環(huán)蛋白,gp43,gage-1,bage-1,psa,muc-1、2、3,和hsp-70,trp-1,gp100/pmel17,β-hcg,ras突變型,p53突變型,hmw黑素瘤抗原,muc-18,hoj-1,細(xì)胞周期蛋白依賴性激酶4(cdk4),半胱天冬酶8,her-2/neu,bcr-abl酪氨酸激酶,癌胚抗原(cea),端粒酶,sv40大t,人乳頭狀瘤病毒hpv6、11、16、18、31和33型;hpv衍生的病毒癌基因e5、e6、e7和l1;存活素、bcl-xl、mcl-1和rho-c。
在本發(fā)明的一個(gè)優(yōu)選實(shí)施方案中,至少一種抗原蛋白質(zhì)或肽或抗原蛋白質(zhì)或肽的片段來自選自下述的癌癥特異性多肽:hpv衍生的病毒癌基因e5、e6、e7和l1;存活素、bcl-xl、mcl-1和rho-c。
與異常生理學(xué)應(yīng)答相關(guān)的抗原
本發(fā)明的一個(gè)實(shí)施方案涉及核酸構(gòu)建體,其中至少一種抗原蛋白質(zhì)或肽或抗原蛋白質(zhì)或肽的片段來自與異常生理學(xué)應(yīng)答相關(guān)的多肽。此類異常生理學(xué)應(yīng)答包括但不限于自身免疫疾病、過敏反應(yīng)、癌癥和先天性疾病。其例子的非詳盡列表包括疾病例如類風(fēng)濕性關(guān)節(jié)炎、全身性紅斑狼瘡、多發(fā)性硬化、銀屑病和crohn氏病。
操作接頭(operativelinker)
本發(fā)明的一個(gè)方面涉及核酸構(gòu)建體,其中在恒定鏈和抗原蛋白質(zhì)或肽或抗原蛋白質(zhì)或肽的片段之間的操作連接是直接連接或由間隔區(qū)介導(dǎo)的連接。術(shù)語操作接頭應(yīng)理解為核苷酸或氨基酸殘基序列,其以保證核酸或多肽的生物加工的方式使核酸構(gòu)建體或嵌合多肽的2個(gè)部分結(jié)合在一起。如果操作接頭是直接連接,那么各自編碼開放讀碼框或開放讀碼框的片段的2個(gè)核酸彼此緊鄰放置,并且從而也在框內(nèi)。如果操作接頭由間隔區(qū)介導(dǎo),那么一系列核苷酸分別插入編碼至少一種恒定鏈和至少一種抗原肽的核苷酸之間。具有間隔區(qū)是在本發(fā)明的范圍內(nèi)的,其中間隔區(qū)僅是以保留開放讀碼框的方式使本發(fā)明的至少2種元件連接的一系列核苷酸,或間隔區(qū)可以編碼如本文下文定義的一種或多種信號(hào)或分離元件。
在一個(gè)特定實(shí)施方案中,本發(fā)明包括操作接頭,其中操作接頭是間隔區(qū)。
在一個(gè)實(shí)施方案中,本發(fā)明包括編碼關(guān)于mhcii類分子的至少一個(gè)輔助表位的間隔區(qū)。輔助表位的例子是免疫原決定簇例如白喉毒素。特別地白喉毒素b片段cooh-末端區(qū)域已顯示在小鼠中是免疫原性的。此外,hsp70部分或整體、以及其他免疫原性肽例如流感病毒、或具有錨定至hlai類和ii類分子的錨定基序的免疫原性序列或肽也可以在核酸構(gòu)建體的間隔區(qū)中編碼。
在另一個(gè)實(shí)施方案中,核酸構(gòu)建體的間隔區(qū)編碼至少一個(gè)蛋白酶切割位點(diǎn)。溶酶體蛋白酶例如組織蛋白酶、天冬氨酸蛋白酶和鋅蛋白酶以及其他細(xì)胞內(nèi)蛋白酶的切割位點(diǎn)落入本發(fā)明的范圍內(nèi)。
在另外一個(gè)實(shí)施方案中,核酸構(gòu)建體的操作接頭可以包括至少一個(gè)sirna或mirna編碼序列。sirna(小干擾rna)和mirna(微小rna)以序列特異性方式靶向內(nèi)源rna用于降解。因此可以選擇在本發(fā)明的核酸構(gòu)建體內(nèi)編碼的sirna或mirna,以靶向不希望有的基因產(chǎn)物。
在另一個(gè)實(shí)施方案中,操作接頭包括至少一個(gè)多聚接頭或多克隆位點(diǎn)(mcs)。多聚接頭和mcs是包括限制酶識(shí)別序列的核苷酸系列,即其中限制酶以平端或交錯(cuò)方式切割dna的位點(diǎn),促進(jìn)dna的其他片段/序列亞克隆到核酸構(gòu)建體內(nèi)。多聚接頭/mcs的識(shí)別序列一般是獨(dú)特的,意味著它們?cè)诤怂針?gòu)建體的其他地方未發(fā)現(xiàn)。操作接頭可以另外包括一個(gè)或多個(gè)終止或末端密碼子,其發(fā)出新生多肽從核糖體釋放的信號(hào)。操作接頭還可以包括至少一個(gè)ires(內(nèi)部核糖體進(jìn)入位點(diǎn))和/或至少一個(gè)啟動(dòng)子。ires是允許在信使rna(mrna)序列中間起始翻譯的核苷酸序列,作為蛋白質(zhì)合成的更大過程的部分。啟動(dòng)子是使得基因能夠被轉(zhuǎn)錄的dna序列。啟動(dòng)子由rna聚合酶識(shí)別,這隨后起始轉(zhuǎn)錄,參見下文。啟動(dòng)子可以是單向或雙向的。
在一個(gè)實(shí)施方案中,跨越在恒定鏈和至少一種抗原之間的區(qū)域的操作接頭是這樣的操作接頭,其包括至少一個(gè)多聚接頭和至少一個(gè)啟動(dòng)子以及還任選地至少一個(gè)ires。這些元件可以以任何次序放置。在一個(gè)進(jìn)一步優(yōu)選的實(shí)施方案中,恒定鏈的終止密碼子已被缺失,并且多聚接頭已以保存開放讀碼框的方式克隆到載體內(nèi),允許插入多聚接頭內(nèi)的至少一種抗原的框內(nèi)閱讀。這具有促進(jìn)多種抗原在一個(gè)步驟或多個(gè)克隆步驟中亞克隆到相同構(gòu)建體內(nèi),并且允許多種抗原在與恒定鏈相同的框內(nèi)同時(shí)表達(dá)的優(yōu)點(diǎn)。終止密碼子可以在多聚接頭后插入用于翻譯終止。這個(gè)實(shí)施方案可以與上述輔助表位中的任何、mi/sirna或本文描述的其他元件中的任何進(jìn)行組合。
本發(fā)明的一個(gè)實(shí)施方案涉及操作接頭相關(guān)于至少一種恒定鏈和至少一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的片段的放置,其中至少一種抗原肽編碼序列放置在:恒定鏈序列內(nèi)、恒定鏈序列的前端、恒定鏈序列的末端部分。這以確保構(gòu)建體的開放讀碼框的可讀性的方式完成,從而使得至少一個(gè)操作接頭在抗原肽之前、周圍或之后。
本發(fā)明的另一個(gè)實(shí)施方案進(jìn)一步涉及操作接頭相關(guān)于至少一種恒定鏈和至少一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的片段的放置,其中至少一種抗原肽編碼序列優(yōu)選放置在恒定鏈的末端部分,并且操作接頭插入兩者之間;末端部分是恒定鏈或其片段的第一個(gè)或最后一個(gè)殘基。
在另一個(gè)實(shí)施方案中,核酸構(gòu)建體不包括操作接頭;相反,至少一種抗原肽編碼序列與恒定鏈直接栓系(tethered)。至少一種抗原肽編碼序列可以放置在:恒定鏈序列內(nèi)、恒定鏈序列的前端、恒定鏈序列的末端部分。這以確保構(gòu)建體的開放讀碼框的可讀性的方式完成。
存在關(guān)于包括或排除操作接頭的2種可能性的優(yōu)點(diǎn);在一個(gè)實(shí)施方案中,排除接頭可以減少引發(fā)針對(duì)所述接頭的免疫應(yīng)答而不是引發(fā)針對(duì)抗原肽的免疫的可能性。
組合
核酸構(gòu)建體編碼多個(gè)元件是在本發(fā)明的范圍內(nèi)的。元件是至少一種恒定鏈或其變體和至少一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的片段。因此具有多個(gè)恒定鏈或其變體落入本發(fā)明的范圍內(nèi),這些各自彼此可操作地連接且與多個(gè)抗原蛋白質(zhì)或肽或抗原蛋白質(zhì)或肽的片段可操作地連接。核酸構(gòu)建體的元件因此必須彼此是可操作地連接的。各自與一種抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的片段可操作地連接的幾個(gè)系列的恒定鏈或其變體(這些系列各自彼此可操作地連接)包含在本發(fā)明內(nèi)。
本發(fā)明的優(yōu)點(diǎn)和非常重要的方面涉及下述事實(shí):可以起始任何類型的免疫應(yīng)答例如t細(xì)胞介導(dǎo)和抗體介導(dǎo)的應(yīng)答,二者可以用已知是弱抗原的表位、未知抗原性質(zhì)的多肽和同時(shí)多個(gè)表位/抗原來起始。
因此,也在本發(fā)明的范圍內(nèi)的是一個(gè)優(yōu)選實(shí)施方案是核酸構(gòu)建體,所述核酸構(gòu)建體編碼與多個(gè)抗原蛋白質(zhì)或肽或蛋白質(zhì)或肽的片段可操作地連接的至少一種恒定鏈或其變體,例如2、3、4、5、6、8、10、12種或更多種抗原蛋白質(zhì)或肽或蛋白質(zhì)或肽的片段。
核酸構(gòu)建體可以包括另外元件。這些包括但不限于:內(nèi)部核糖體進(jìn)入位點(diǎn)(ires);編碼涉及抗原呈遞的蛋白質(zhì)的基因,所述蛋白質(zhì)例如lamp、鈣網(wǎng)織蛋白、hsp33、hsp60、hsp70、hsp90、hsp100、shsp(小熱休克蛋白)和熱休克結(jié)合蛋白例如77-殘基dnaj-同源hsp73-結(jié)合結(jié)構(gòu)域;編碼涉及細(xì)胞內(nèi)散布(spreading)的蛋白質(zhì)的基因,所述蛋白質(zhì)例如vp22、hivtat、cx43或其他連接蛋白和細(xì)胞內(nèi)縫隙連接組成成分;編碼天然殺傷細(xì)胞(nk細(xì)胞)活化分子例如h60和細(xì)胞因子、雞卵白蛋白或任何t輔助細(xì)胞表位的基因。
在本發(fā)明的一個(gè)優(yōu)選實(shí)施方案中,核酸構(gòu)建體包括編碼涉及抗原呈遞的蛋白質(zhì)的至少一種基因,所述蛋白質(zhì)例如lamp、limp、鈣網(wǎng)織蛋白hsp33、hsp60、hsp70、hsp90、hsp100、shsp(小熱休克蛋白)或77-殘基dnaj-同源hsp73-結(jié)合結(jié)構(gòu)域。
在本發(fā)明的另外一個(gè)優(yōu)選實(shí)施方案中,核酸構(gòu)建體包括編碼涉及細(xì)胞內(nèi)散布的蛋白質(zhì)的至少一種基因,所述蛋白質(zhì)例如vp22、cx43、hivtat、其他連接蛋白和細(xì)胞內(nèi)縫隙連接組成成分。
啟動(dòng)子
術(shù)語啟動(dòng)子將在此處用于指一組轉(zhuǎn)錄控制模塊,其在rna聚合酶ii的起始位點(diǎn)周圍成簇。關(guān)于啟動(dòng)子被如何組織的許多想法衍生自幾種病毒啟動(dòng)子的分析,包括關(guān)于hsv胸苷激酶(tk)和sv40早期轉(zhuǎn)錄單位的啟動(dòng)子。由更近期工作加強(qiáng)的這些研究已顯示啟動(dòng)子由不連續(xù)功能模塊組成,各自由約7-20bpdna組成,并且包含轉(zhuǎn)錄激活蛋白質(zhì)的一個(gè)或多個(gè)識(shí)別位點(diǎn)。每個(gè)啟動(dòng)子中的至少一個(gè)模塊作用于rna合成的起始位點(diǎn)的定位。這點(diǎn)的最佳已知例子是tata盒,但在缺乏tata盒的某些啟動(dòng)子中,例如哺乳動(dòng)物末端脫氧核苷酰轉(zhuǎn)移酶基因的啟動(dòng)子和sv40晚期基因的啟動(dòng)子,重疊自身起始位點(diǎn)的不連續(xù)元件幫助固定起始位置。
另外的啟動(dòng)子元件調(diào)節(jié)轉(zhuǎn)錄起始頻率。一般地,這些位于起始位點(diǎn)上游30-110bp的區(qū)域中,盡管許多啟動(dòng)子近期已顯示也包含起始位點(diǎn)下游的功能元件。元件之間的間距是彈性的,從而使得當(dāng)元件倒轉(zhuǎn)或相對(duì)于彼此移動(dòng)時(shí),啟動(dòng)子功能被保存。在tk啟動(dòng)子中,在活性開始下降前元件之間的間距可以增至相隔50bp。依賴于啟動(dòng)子,看起來個(gè)別元件可以共同或獨(dú)立地作用,以激活轉(zhuǎn)錄。可以引導(dǎo)由核酸構(gòu)建體編碼的序列的轉(zhuǎn)錄起始的任何啟動(dòng)子可以在本發(fā)明中使用。
本發(fā)明的一個(gè)方面包括核酸構(gòu)建體,其中使得構(gòu)建體能夠表達(dá)的啟動(dòng)子在至少一種可操作地連接的恒定鏈和抗原蛋白質(zhì)或肽編碼序列前。
進(jìn)一步方面的是啟動(dòng)子選自組成型啟動(dòng)子、誘導(dǎo)型啟動(dòng)子、生物特異性啟動(dòng)子、組織特異性啟動(dòng)子、細(xì)胞類型特異性啟動(dòng)子和炎癥特異性啟動(dòng)子。
啟動(dòng)子的例子包括但不限于:組成型啟動(dòng)子例如:猴病毒40(sv40)早期啟動(dòng)子、小鼠乳腺腫瘤病毒啟動(dòng)子、人免疫缺陷病毒長末端重復(fù)啟動(dòng)子、莫洛尼病毒啟動(dòng)子、禽白血病病毒啟動(dòng)子、eb病毒即早期啟動(dòng)子、勞斯肉瘤病毒(rsv)啟動(dòng)子、人肌動(dòng)蛋白啟動(dòng)子、人肌球蛋白啟動(dòng)子、人血紅蛋白啟動(dòng)子、巨細(xì)胞病毒(cmv)啟動(dòng)子和人肌肉肌酸啟動(dòng)子,誘導(dǎo)型啟動(dòng)子例如:金屬硫蛋白啟動(dòng)子、糖皮質(zhì)激素啟動(dòng)子、孕酮啟動(dòng)子和四環(huán)素啟動(dòng)子(tet開或tet關(guān)),組織特異性啟動(dòng)子例如:her-2啟動(dòng)子和psa相關(guān)啟動(dòng)子和雙向啟動(dòng)子,其能夠起始從啟動(dòng)子任一方向的轉(zhuǎn)錄。
使用誘導(dǎo)型啟動(dòng)子的優(yōu)點(diǎn)包括提供可以隨意激活的“休眠”核酸構(gòu)建體的選項(xiàng)。如果免疫應(yīng)答的引發(fā)優(yōu)選僅在體內(nèi)局部與全身性(例如在涉及癌癥的情況下)比較誘導(dǎo),或免疫應(yīng)答的引發(fā)在施用時(shí)對(duì)于接受者的健康是有害的,這可以是有用的。
在一個(gè)優(yōu)選實(shí)施方案中,核酸構(gòu)建體包括選自下述的啟動(dòng)子:cmv啟動(dòng)子、sv40啟動(dòng)子和rsv啟動(dòng)子。
遞送媒介物
本發(fā)明的一個(gè)方面包括如包括在遞送媒介物中的在上文任何中描述的核酸構(gòu)建體。遞送媒介物是由此核苷酸序列或多肽或兩者可以從至少一種介質(zhì)轉(zhuǎn)運(yùn)到另一種介質(zhì)的實(shí)體。遞送媒介物一般用于表達(dá)在核酸構(gòu)建體內(nèi)編碼的序列和/或用于構(gòu)建體或其中編碼的多肽的細(xì)胞內(nèi)遞送。
核酸構(gòu)建體可以在體內(nèi)或先體外后體內(nèi)轉(zhuǎn)移到細(xì)胞內(nèi);后者通過從患者中取出靶組織(即肝細(xì)胞或白血細(xì)胞),在體外轉(zhuǎn)移構(gòu)建體,并且隨后將轉(zhuǎn)導(dǎo)細(xì)胞重新植入患者內(nèi)來進(jìn)行。
非病毒遞送的方法包括物理(無載體遞送)和化學(xué)方法(基于合成載體的遞送)。
物理方法包括針注射、基因槍、射流注射、電穿孔、超聲和水動(dòng)力遞送,采用滲透細(xì)胞膜且促進(jìn)細(xì)胞內(nèi)基因轉(zhuǎn)移的物理力。所述物理力可以是電或機(jī)械的。
化學(xué)方法使用合成或天然存在的化合物作為載體,以將轉(zhuǎn)基因遞送到細(xì)胞內(nèi)。用于非病毒基因遞送的最頻繁研究的策略是通過使用陽離子脂質(zhì)或陽離子聚合物將dna配制成濃縮顆粒。含dna的顆粒隨后經(jīng)由以細(xì)胞內(nèi)囊泡形式的胞吞作用、巨胞飲作用或吞噬作用被細(xì)胞吸收,dna的小部分從細(xì)胞內(nèi)囊泡中釋放到細(xì)胞質(zhì)內(nèi)并移動(dòng)到核內(nèi),在其中發(fā)生轉(zhuǎn)基因表達(dá)。
在本發(fā)明的范圍內(nèi)的是遞送媒介物是選自下述的媒介物:基于rna的媒介物、基于dna的媒介物/載體、基于脂質(zhì)的媒介物、基于聚合物的媒介物和病毒衍生的dna或rna媒介物。
本發(fā)明的一個(gè)優(yōu)選實(shí)施方案涉及通過機(jī)械或電技術(shù)遞送核酸構(gòu)建體。
?物理
○注射:一個(gè)優(yōu)選實(shí)施方案涉及在溶液中的核酸構(gòu)建體的簡單注射。注射通常用遞送至細(xì)胞外空間的核酸構(gòu)建體在骨骼肌中肌內(nèi)(im)、皮內(nèi)(id)或?qū)τ诟芜M(jìn)行。通過注射的遞送可以通過下述得到輔助:電穿孔;使用鹽水或蔗糖的高滲溶液;用肌毒素例如布比卡因暫時(shí)損害肌纖維;或加入能夠增強(qiáng)dna被靶細(xì)胞內(nèi)在化效率的物質(zhì),例如轉(zhuǎn)鐵蛋白、與水不混溶的溶劑、非離子聚合物、表面活性劑或核酸酶抑制劑。
○基因槍:基因槍或biolistic粒子遞送系統(tǒng)是用于將遺傳信息注射細(xì)胞的裝置。有效負(fù)荷(payload)是用例如質(zhì)粒dna包被的重金屬例如金、銀或鎢的元素粒子。這種技術(shù)通常簡稱為biolistics。壓縮氦可以用作推進(jìn)劑。特別地在金顆粒例如膠體金顆粒上的核酸構(gòu)建體涂層是一個(gè)有利的實(shí)施方案。
○氣動(dòng)(射流)注射:不需要顆粒,水性溶液。
○電穿孔或電滲透作用是由外部應(yīng)用的電場引起的細(xì)胞質(zhì)膜的導(dǎo)電性和滲透性的顯著增加。
○超聲促進(jìn)的基因轉(zhuǎn)移:超聲產(chǎn)生膜孔,并且促進(jìn)通過dna被動(dòng)擴(kuò)散跨越膜孔的細(xì)胞內(nèi)基因轉(zhuǎn)移。效率可以通過使用造影劑或使得膜更具有流動(dòng)性的條件得到增強(qiáng)。
○水動(dòng)力基因遞送:水動(dòng)力基因遞送是將裸露質(zhì)粒dna引入在高度充滿(perfused)的內(nèi)臟(例如肝)的細(xì)胞內(nèi)的簡單方法。在嚙齒類動(dòng)物中,大量dna溶液的快速尾靜脈注射引起注射溶液在下腔靜脈的暫時(shí)溢流,這超過心輸出量。因此,注射誘導(dǎo)dna溶液逆流到肝內(nèi)的流動(dòng)、肝內(nèi)壓力的快速升高、肝擴(kuò)大和肝窗的可逆破裂。
?化學(xué)
○陽離子脂質(zhì)介導(dǎo)的基因遞送:盡管某些陽離子脂質(zhì)單獨(dú)顯示出良好的轉(zhuǎn)染活性,但它們通常用不荷電磷脂或膽固醇作為輔助脂質(zhì)進(jìn)行配制,以形成脂質(zhì)體。在與陽離子脂質(zhì)體混合后,使質(zhì)粒dna濃縮成稱為lipoplex的準(zhǔn)穩(wěn)定顆粒。lipoplex中的dna被充分保護(hù)免于核酸酶降解。lipoplex能夠觸發(fā)細(xì)胞攝取,并且促進(jìn)在到達(dá)破壞性溶酶體區(qū)室前使dna從細(xì)胞內(nèi)囊泡中釋放。
○陽離子聚合物介導(dǎo)的基因遞送:大多數(shù)陽離子聚合物都具有使dna濃縮成為小顆粒并且通過與細(xì)胞表面上陰離子位點(diǎn)的電荷-電荷相互作用促進(jìn)經(jīng)由胞吞作用的細(xì)胞攝取的功能。陽離子聚合物dna載體包括聚乙烯亞胺(pei)、聚乙二胺和聚丙胺樹狀聚體(dendrimers)、聚烯丙胺、陽離子右旋糖酐、殼聚糖、陽離子蛋白質(zhì)(聚賴氨酸、魚精蛋白和組蛋白)和陽離子肽。
○脂質(zhì)-聚合物雜交系統(tǒng):用聚陽離子預(yù)濃縮dna,隨后用陽離子脂質(zhì)體、陰離子脂質(zhì)體或兩親性聚合物連同或不連同輔助脂質(zhì)包被。
化學(xué)遞送媒介物的例子包括但不限于:生物可降解聚合物微球體、基于脂質(zhì)的制劑例如脂質(zhì)體載體、陽離子荷電分子例如脂質(zhì)體、鈣鹽或樹狀聚體、脂多糖、多肽和多糖。
可選的物理遞送方法可以包括裸露核酸構(gòu)建體在粘膜表面例如鼻和肺粘膜上的氣溶膠滴注;核酸構(gòu)建體向眼和粘膜組織的局部施用;和水合作用例如通過其鹽水溶液受力進(jìn)入眼的角膜基質(zhì)內(nèi)的基質(zhì)水合。
本發(fā)明的另一個(gè)實(shí)施方案包括載體,其在本文中指示病毒載體(即非病毒),作為遞送媒介物。根據(jù)本發(fā)明的病毒載體由修飾的病毒基因組制成,即形成病毒基因組的實(shí)際dna或rna,并且以裸露形式引入。因此,在由病毒或非病毒蛋白質(zhì)制成的病毒基因組周圍的任何包被結(jié)構(gòu)不是根據(jù)本發(fā)明的病毒載體的部分。
病毒載體由其衍生的病毒選自下述非詳盡組:腺病毒、逆轉(zhuǎn)錄病毒、慢病毒、腺伴隨病毒、皰疹病毒、痘苗病毒、泡沫病毒、巨細(xì)胞病毒、塞姆利基森林病毒、痘病毒、rna病毒載體和dna病毒載體。此類病毒載體是本領(lǐng)域眾所周知的。
重組細(xì)胞
本發(fā)明的一個(gè)方面涉及包括如在上文中的任何中定義的核酸構(gòu)建體的細(xì)胞。此類重組細(xì)胞可以用作體外研究的工具,用作核酸構(gòu)建體的遞送媒介物或用作基因療法方案的一部分。根據(jù)本發(fā)明的核酸構(gòu)建體可以通過本領(lǐng)域眾所周知的技術(shù)引入細(xì)胞內(nèi),并且所述技術(shù)包括dna顯微注射到細(xì)胞的核內(nèi)、轉(zhuǎn)染、電穿孔、脂轉(zhuǎn)染/脂質(zhì)體融合和粒子轟擊。合適的細(xì)胞包括自體和非自體細(xì)胞,并且可以包括異種細(xì)胞。
在一個(gè)優(yōu)選實(shí)施方案中,本發(fā)明的核酸構(gòu)建體包括在抗原呈遞細(xì)胞(apc)內(nèi)。在其表面上呈遞與mhc分子結(jié)合的抗原的任何細(xì)胞被視為抗原呈遞細(xì)胞。此類細(xì)胞包括但不限于巨噬細(xì)胞、樹突細(xì)胞、b細(xì)胞、雜合apc和養(yǎng)育(foster)apc。制備雜合apc的方法是本領(lǐng)域眾所周知的。
在一個(gè)更優(yōu)選的實(shí)施方案中,apc是專職抗原呈遞細(xì)胞,并且最優(yōu)選apc是mhc-i和/或mhc-ii表達(dá)細(xì)胞。
根據(jù)上文中的任何的apc可以是得自患者的干細(xì)胞。在引入本發(fā)明的核酸構(gòu)建體后,干細(xì)胞可以以治療具有醫(yī)學(xué)狀況的患者的意圖重新引入患者內(nèi)。優(yōu)選地,從患者中分離的細(xì)胞是能夠分化成抗原呈遞細(xì)胞的干細(xì)胞。
在本發(fā)明的范圍內(nèi)另外包括的是包括本發(fā)明的核酸構(gòu)建體的抗原呈遞細(xì)胞不表達(dá)任何共刺激信號(hào),并且抗原蛋白質(zhì)或肽或所述蛋白質(zhì)或肽的抗原片段是自身抗原。
嵌合蛋白質(zhì)和抗體
本發(fā)明的一個(gè)目的是由如本文上文描述的核酸構(gòu)建體編碼的嵌合蛋白質(zhì),包括至少一種可操作地連接的恒定鏈或其變體和至少一種抗原蛋白質(zhì)或肽或所述抗原蛋白質(zhì)或肽的片段。嵌合蛋白質(zhì)應(yīng)理解為由核苷酸序列編碼的遺傳改造蛋白質(zhì),所述核苷酸序列通過將2種或更多種完全或部分基因或一系列(非)隨機(jī)核酸剪接在一起制備。
本發(fā)明的一個(gè)方面涉及可以識(shí)別如本文上文定義的嵌合蛋白質(zhì)的抗體。術(shù)語抗體應(yīng)理解為免疫球蛋白分子和免疫球蛋白分子的活性部分。抗體是例如完整免疫球蛋白分子或其保留免疫學(xué)活性的片段。此類抗體可以用于動(dòng)物的被動(dòng)免疫接種,或用于在用于檢測抗體與之結(jié)合的蛋白質(zhì)的測定中。
核酸構(gòu)建體組合物
本發(fā)明的一個(gè)方面涉及包括核酸序列的組合物,所述核酸序列編碼與至少一種抗原蛋白質(zhì)或肽或所述抗原蛋白質(zhì)或肽的片段可操作地連接的至少一種恒定鏈或其變體。組合物因此可以包括如上文中的任何中定義的核酸構(gòu)建體。組合物可以另外用作藥物。
根據(jù)本發(fā)明的核酸構(gòu)建體組合物可以根據(jù)已知方法進(jìn)行配制,例如通過一種或多種藥學(xué)上可接受的載體也稱為賦形劑或穩(wěn)定劑與活性劑的混合。這些賦形劑對(duì)于施用于任何個(gè)體/動(dòng)物,優(yōu)選脊椎動(dòng)物且更優(yōu)選人可以是可接受的,因?yàn)樗鼈冊(cè)诓捎玫膭┝亢蜐舛认聦?duì)暴露于其的細(xì)胞或個(gè)體是無毒的。通常生理學(xué)上可接受的載體是ph緩沖水溶液。此類賦形劑、載體和配制方法的例子可以例如在remington'spharmaceuticalsciences(maackpublishingco,easton,pa)中找到。生理學(xué)上可接受的載體的例子包括但不限于:緩沖液例如磷酸鹽、檸檬酸鹽和其他有機(jī)酸;抗氧化劑包括抗壞血酸;低分子量(小于約10個(gè)殘基)多肽;蛋白質(zhì)例如血清白蛋白、明膠或免疫球蛋白;親水聚合物例如聚乙烯吡咯烷酮;氨基酸例如甘氨酸、谷氨酰胺、天冬酰胺、精氨酸或賴氨酸;單糖、二糖和其他碳水化合物包括葡萄糖、甘露糖或糊精;螯合劑例如edta;糖醇例如甘露糖醇或山梨糖醇;成鹽抗衡離子例如鈉;和/或非離子表面活性劑例如tweentm、聚乙二醇(peg)和pluronicstm。
為了配制適合于有效施用的藥學(xué)上可接受的組合物,此類組合物根據(jù)本發(fā)明將包含有效量的核酸構(gòu)建體,所述核酸構(gòu)建體被包括在遞送媒介物內(nèi)或嵌合蛋白質(zhì)在如本文描述的核酸構(gòu)建體內(nèi)編碼。通常,如果用如由本發(fā)明的核酸構(gòu)建體編碼的蛋白質(zhì)或多肽引發(fā)免疫應(yīng)答,那么通過使蛋白質(zhì)或肽與之偶聯(lián)且因此幫助誘導(dǎo)免疫應(yīng)答,載體將用作支架。載體蛋白質(zhì)可以是任何常規(guī)載體,包括適合于呈遞免疫原決定簇的任何蛋白質(zhì)。合適載體一般是大的、緩慢代謝的大分子,例如蛋白質(zhì)、多糖、聚乳酸、聚乙醇酸、聚合氨基酸、氨基酸共聚物、脂質(zhì)聚集體(例如油小滴或脂質(zhì)體)和非活性病毒顆粒。此類載體是本領(lǐng)域普通技術(shù)人員眾所周知的。此外,這些載體可以充當(dāng)免疫刺激劑(“佐劑”)。動(dòng)物的免疫接種可以用佐劑和/或藥物載體執(zhí)行。常規(guī)載體蛋白質(zhì)包括但不限于鑰孔血藍(lán)蛋白,血清蛋白質(zhì)例如轉(zhuǎn)鐵蛋白、牛血清白蛋白或人血清白蛋白,卵白蛋白,免疫球蛋白或激素例如胰島素。載體可以連同佐劑或獨(dú)立于其存在。
在下文中,核酸構(gòu)建體組合物或組合物意指包含對(duì)于預(yù)防和治療用途包括刺激患者中的免疫應(yīng)答有用的組合物。進(jìn)一步考慮本發(fā)明的組合物不包括任何全身或局部毒性反應(yīng)或任何其他副作用。
在一個(gè)優(yōu)選實(shí)施方案中,如本文使用的,短語‘組合物’指用于引發(fā)免疫應(yīng)答的組合物。
在一個(gè)優(yōu)選實(shí)施方案中,核酸構(gòu)建體是包裝的。用于核酸構(gòu)建體的包裝裝置(means)包括選自但不限于下述的裝置:基于rna的或基于dna載體、基于脂質(zhì)的載體、病毒表達(dá)載體、病毒遞送載體、膠體金顆粒和生物可降解聚合物微球體的包被。先前提及的遞送裝置中的任何因此可以用于在組合物中使用的包裝目的。
在一個(gè)實(shí)施方案中,核酸構(gòu)建體的包裝方法是選自但不限于下述的病毒表達(dá)載體:腺病毒、逆轉(zhuǎn)錄病毒、慢病毒、腺伴隨病毒、皰疹病毒、痘苗病毒和dna病毒載體。病毒載體可以是復(fù)制缺陷型或條件復(fù)制缺陷型病毒載體。
本發(fā)明的一個(gè)方面涉及包括至少2種載體的組合物。這包含如所述的任何1種或2種不同核酸構(gòu)建體可以包裝到至少2種載體內(nèi),這些載體是如上文中的任何中所述的類型。本發(fā)明另外涉及包括3、4、5或6種載體的組合物。再次,這些載體可以彼此不同或相同,并且可以攜帶如本文上文描述的等同或不同核酸構(gòu)建體。本發(fā)明的一個(gè)進(jìn)一步方面涉及組合物,其包括如由本文描述的核酸構(gòu)建體中的任何編碼的至少一種嵌合蛋白質(zhì)。當(dāng)嵌合蛋白質(zhì)或多肽待用作免疫原時(shí),它可以通過在重組細(xì)胞中表達(dá)任何一種或多種上述核酸構(gòu)建體產(chǎn)生,或它可以通過本領(lǐng)域已知的方法通過化學(xué)合成制備。如上文中所述,此類嵌合蛋白質(zhì)和/或肽可以與載體偶聯(lián),以增加對(duì)于蛋白質(zhì)/肽的免疫應(yīng)答,并且可以連同或不連同佐劑和/或賦形劑一起施用。
在一個(gè)實(shí)施方案中,本發(fā)明涉及如本文描述的核酸構(gòu)建體用于產(chǎn)生組合物的用途。
增強(qiáng)免疫應(yīng)答:常規(guī)佐劑
佐劑可以包括在組合物中以增強(qiáng)特異性免疫應(yīng)答。因此,特別重要的是鑒定這樣的佐劑,當(dāng)與一種或多種抗原/核酸構(gòu)建體和/或遞送媒介物(其中任何也可以被稱為免疫原決定簇)組合時(shí),其導(dǎo)致能夠誘導(dǎo)強(qiáng)特異性免疫應(yīng)答的組合物。免疫原決定簇還可以在免疫接種前與2種或更多種不同佐劑混合。組合物在呈現(xiàn)的正文中還被稱為免疫原性組合物。
大量佐劑已得到描述并且用于在實(shí)驗(yàn)室動(dòng)物例如小鼠、大鼠和兔中生成抗體。在此類設(shè)置中,副作用的耐受相當(dāng)高,因?yàn)橹饕繕?biāo)是獲得強(qiáng)抗體應(yīng)答。對(duì)于用途和對(duì)于批準(zhǔn)用于在藥物中使用,且特別對(duì)于在人中使用,要求組合物的組分包括佐劑是充分表征的。進(jìn)一步要求組合物具有任何不良反應(yīng)的最低限度危險(xiǎn),例如肉芽腫、膿腫或發(fā)熱。
本發(fā)明的一個(gè)實(shí)施方案涉及包括佐劑的組合物。在一個(gè)優(yōu)選實(shí)施方案中,組合物適合于施用于哺乳動(dòng)物例如人類。因此,優(yōu)選佐劑適合于施用于哺乳動(dòng)物,并且最優(yōu)選地適合于施用于人類。
在另一個(gè)優(yōu)選實(shí)施方案中,組合物適合于施用于鳥或魚,并且最優(yōu)選雞(家雞)。因此,優(yōu)選佐劑適合于施用于鳥或魚。
佐劑的選擇可以進(jìn)一步通過其刺激希望的免疫應(yīng)答類型b細(xì)胞和/或t細(xì)胞激活的能力進(jìn)行選擇,并且組合物可以配制為最優(yōu)化對(duì)相關(guān)淋巴組織的分布和呈遞。
關(guān)于本發(fā)明的佐劑可以根據(jù)其起源進(jìn)行分組,其為礦物質(zhì)、細(xì)菌、植物、合成的或宿主產(chǎn)物。在這個(gè)分類法下的第一個(gè)組是礦物質(zhì)佐劑,例如鋁化合物。與鋁鹽沉淀的抗原或與預(yù)形成的鋁鹽或化合物混合或吸附的抗原已廣泛用于增強(qiáng)動(dòng)物和人中的免疫應(yīng)答。鋁顆粒已在免疫接種后7天在兔的局部淋巴結(jié)中得到證實(shí),并且可能另一種顯著功能是將抗原導(dǎo)向在結(jié)自身中的含t細(xì)胞區(qū)域。佐劑效力已顯示與引流淋巴結(jié)的暗示相關(guān)。雖然許多研究已證實(shí)與鋁鹽一起施用的抗原導(dǎo)致增加的體液免疫,細(xì)胞介導(dǎo)的免疫看起來僅是輕微增加的,如通過遲發(fā)型超敏反應(yīng)測量的。氫氧化鋁已描述為激活補(bǔ)體途徑。這種機(jī)制可以在局部炎癥應(yīng)答以及免疫球蛋白生產(chǎn)和b細(xì)胞記憶中起作用。此外,氫氧化鋁可以保護(hù)抗原不受快速分解代謝。主要由于其極佳的安全記錄,鋁化合物是目前在人中使用的唯一佐劑。
另一大組佐劑是細(xì)菌起源的那些。具有細(xì)菌起源的佐劑可以進(jìn)行純化且合成(例如胞壁酰二肽、脂質(zhì)a),并且宿主介質(zhì)已得到克隆(白介素1和2)。最近十年已在細(xì)菌起源活性組分的幾種佐劑的化學(xué)純化中獲得顯著進(jìn)展:百日咳博德特氏菌、結(jié)核分枝桿菌、脂多糖、弗氏完全佐劑(fca)和弗氏不完全佐劑(difcolaboratories,detroit,mich.)和merckadjuvant65(merck和company,inc.,rahway,n.j.)。另外,依照本發(fā)明的合適佐劑是例如titermaxclassical佐劑(sigma-aldrich)、iscoms、quila、alun,參見us58767和5,554,372,脂質(zhì)a衍生物、霍亂毒素衍生物、hsp衍生物、lps衍生物、合成肽基質(zhì)、gmdp及其他以及與免疫刺激劑組合(us5,876,735)。百日咳博德特氏菌在本發(fā)明的背景中作為佐劑是有利的,由于其通過對(duì)t淋巴細(xì)胞群體的作用調(diào)節(jié)細(xì)胞介導(dǎo)的免疫的能力。對(duì)于脂多糖和弗氏完全佐劑,佐劑活性部分已得到鑒定且合成,這允許研究結(jié)構(gòu)-功能關(guān)系。這些還考慮用于包括在根據(jù)本發(fā)明的免疫原性組合物中。
已發(fā)現(xiàn)脂多糖(lps)及其各種衍生物包括脂質(zhì)a與脂質(zhì)體或其他脂質(zhì)乳劑組合是有力佐劑。仍未確定是否可以產(chǎn)生用于在人中的一般使用的具有足夠低毒性的衍生物。弗氏完全佐劑是大多數(shù)實(shí)驗(yàn)研究中的標(biāo)準(zhǔn)。
礦物油可以加入免疫原性組合物中,以便保護(hù)抗原不受快速分解代謝。
許多其他類型的材料可以用作根據(jù)本發(fā)明的免疫原性組合物中的佐劑。這些包括植物產(chǎn)物例如皂苷、動(dòng)物產(chǎn)物例如殼多糖和眾多合成化學(xué)制品。
根據(jù)本發(fā)明的佐劑還可以通過其提議的作用機(jī)制進(jìn)行分類。這類分類法必定是略微任意的,因?yàn)榇蠖鄶?shù)佐劑看起來通過超過一種機(jī)制起作用。佐劑可以通過抗原定位和遞送,或通過對(duì)構(gòu)成免疫系統(tǒng)的細(xì)胞例如巨噬細(xì)胞和淋巴細(xì)胞的直接作用起作用。根據(jù)本發(fā)明的佐劑通過其增強(qiáng)免疫應(yīng)答的另一種機(jī)制是通過抗原沉積物的產(chǎn)生。這看起來促成鋁化合物、油乳劑、脂質(zhì)體和合成聚合物的佐劑活性。脂多糖和胞壁酰二肽的佐劑活性看起來主要通過巨噬細(xì)胞的激活介導(dǎo),而百日咳博德特氏菌影響巨噬細(xì)胞和淋巴細(xì)胞。當(dāng)摻入根據(jù)本發(fā)明的免疫原性組合物內(nèi)可以是有用的佐劑的進(jìn)一步例子在us5,554,372中描述。
在根據(jù)本發(fā)明的組合物中有用的佐劑因此可以是礦物鹽,例如氫氧化鋁和磷酸鋁或鈣凝膠、油乳劑和基于表面活性劑的制劑例如mf59(微流體化去污劑穩(wěn)定的水包油乳劑)、qs21(純化皂苷)、as02(sbas2、水包油乳劑+單磷酰脂質(zhì)a(mpl)+qs21)、montanideisa51和isa-720(穩(wěn)定的油包水乳劑)、佐劑65(包含花生油、二縮甘露醇一油酸和單硬脂酸鋁)、ribiimmunochemresearchinc.,hamilton,utah)、微粒佐劑例如病毒體(摻入流感血凝素的單層脂質(zhì)體小泡(原文為cehicles,疑為vehicles))、as04(含mpl的al鹽)、iscoms(皂苷和脂質(zhì)(例如膽固醇)的結(jié)構(gòu)復(fù)合物、聚丙交酯共乙交酯(plg)、微生物衍生物(天然和合成)例如單磷酰脂質(zhì)a(mpl)、detox(mpl+草分支桿菌(m.phlei)細(xì)胞壁支架)、agp(rc-529(合成乙酰化單糖))、dc_chol(能夠自組構(gòu)成脂質(zhì)體的類脂免疫刺激劑)、om-174(脂質(zhì)a衍生物)、cpg基序(包含免疫刺激cpg基序的合成寡核苷酸)、修飾細(xì)菌毒素、具有無毒佐劑效應(yīng)的lt和ct、內(nèi)源人免疫調(diào)節(jié)劑例如hgm-csf或hil-12或immudaptin(c3d串聯(lián)排列)、惰性媒介物例如金顆粒。
佐劑的另外例子包括:免疫刺激油乳劑(例如,油包水、水包油、水包油包水例如弗氏不完全佐劑例如montainde?,specol,礦物鹽例如al(oh)3、alpo4,微生物產(chǎn)物,皂苷例如quala,合成產(chǎn)物以及佐劑佐劑,和免疫刺激復(fù)合物(iscom)和細(xì)胞因子,熱滅活細(xì)菌/組分,納米珠,lps,lta。其他常用佐劑的列表公開于wo2003/089471中的第6-8頁上,所述列表在此引入作為參考。
根據(jù)本發(fā)明的免疫原性組合物還可以包含稀釋劑例如緩沖劑,抗氧化劑例如抗壞血酸,低分子量(小于約10個(gè)殘基)多肽,蛋白質(zhì),氨基酸,碳水化合物包括葡萄糖、甘露糖或糊精,螯合劑例如edta,谷胱甘肽及其他穩(wěn)定劑和賦形劑。中性緩沖鹽水或與非特異性血清白蛋白混合的鹽水是示例性合適稀釋劑。
佐劑一般以根據(jù)制造商的說明書的量包括在免疫原性組合物中。
增強(qiáng)免疫應(yīng)答:非常規(guī)佐劑
細(xì)胞因子調(diào)節(jié)
對(duì)于有效的疫苗,它必須誘導(dǎo)對(duì)于給定病原體的合適免疫應(yīng)答。這可以通過修飾表達(dá)的抗原形式(即細(xì)胞內(nèi)與分泌的比較)、遞送方法和途徑和遞送的dna劑量來完成。然而,它還可以通過共施用編碼免疫調(diào)節(jié)分子例如細(xì)胞因子、淋巴因子或共刺激分子的質(zhì)粒dna(pdna)來完成。這些“遺傳佐劑”可以連同如本文概述的‘常規(guī)佐劑’或‘其他免疫刺激佐劑’中的任何以許多方式施用:
?作為2種分開質(zhì)粒的混合物,一種編碼免疫原和另一種編碼細(xì)胞因子;
?作為通過間隔區(qū)分開的單一的雙或多順反子載體;或
?作為質(zhì)粒編碼的嵌合體或融合蛋白;或
?以其天然形式,即蛋白質(zhì)或核苷酸。
一般而言,促炎因子(例如各種白介素、腫瘤壞死因子和gm-csf)加上th2誘導(dǎo)細(xì)胞因子的共施用增加抗體應(yīng)答,而促炎因子和th1誘導(dǎo)細(xì)胞因子降低體液應(yīng)答且增加細(xì)胞毒性應(yīng)答(這例如在病毒保護(hù)中更重要)。有時(shí)還使用共刺激分子如b7-1、b7-2和cd40l。
這個(gè)概念已成功地應(yīng)用于編碼il-10的pdna的局部施用。編碼b7-1(在apc上的配體)的質(zhì)粒在抗腫瘤模型中已成功地增強(qiáng)免疫應(yīng)答,并且使編碼gm-csf和約氏瘧原蟲(p.yoelii)的環(huán)子孢子蛋白(pycsp)的質(zhì)粒混合已增強(qiáng)針對(duì)后續(xù)攻擊的保護(hù)(而單獨(dú)的編碼pycsp的質(zhì)粒則不是)。gm-csf可以促使樹突細(xì)胞更有效地呈遞抗原,且增強(qiáng)il-2產(chǎn)生和th細(xì)胞激活,從而驅(qū)動(dòng)增加的免疫應(yīng)答。這可以通過首先用ppycsp和pgm-csf混合物引發(fā),且以后用表達(dá)pycsp的重組痘病毒加強(qiáng)得到進(jìn)一步增強(qiáng)。然而,與通過單獨(dú)的ppcmsp1-hbs遞送獲得的保護(hù)比較,編碼gm-csf(或ifn-γ、或il-2)和夏氏瘧原蟲(p.chabaudi)裂殖子表面蛋白質(zhì)1(c末端)-乙型肝炎病毒表面蛋白質(zhì)(pcmsp1-hbs)的融合蛋白的質(zhì)粒的共注射實(shí)際上消除了針對(duì)攻擊的保護(hù)。
其他免疫刺激佐劑
在一個(gè)實(shí)施方案中,下述中的任何可以用作關(guān)于根據(jù)本發(fā)明的核酸構(gòu)建體或組合物的免疫刺激佐劑:
lps(脂多糖),poly-ic(多聚肌醇胞嘧啶)或任何其他佐劑,其類似于雙鏈rna、ll37、rig-1解螺旋酶、il-12、il-18、ccl-1、ccl-5、ccl-19、ccl-21、gm-csf、cx3cl、cd86、pd-1、分泌pd-1、il10-r、分泌il10-r、il21、icosl、41bbl、cd40l,以及刺激免疫應(yīng)答的其他任何蛋白質(zhì)或核酸序列。
在一個(gè)實(shí)施方案中,使免疫刺激佐劑與腺病毒纖維蛋白質(zhì)融合。例如,cx3cl可以與腺病毒纖維蛋白質(zhì)融合。
免疫刺激cpg基序
質(zhì)粒dna自身看起來對(duì)免疫系統(tǒng)具有佐劑作用。質(zhì)粒dna已衍生自這樣的細(xì)菌,發(fā)現(xiàn)所述細(xì)菌觸發(fā)先天性免疫防御機(jī)制,激活樹突細(xì)胞和產(chǎn)生th1細(xì)胞因子。這是由于識(shí)別具有免疫刺激的的特定cpg二核苷酸序列。cpg刺激(cpg-s)序列在細(xì)菌衍生的dna中的出現(xiàn)頻率為真核生物中的20倍。這是因?yàn)檎婧松镲@示“cpg抑制”-即cpg二核苷酸對(duì)以比預(yù)期的低得多的頻率出現(xiàn)。另外,cpg-s序列是低甲基化的。這在細(xì)菌dna中頻繁出現(xiàn),而在真核生物中出現(xiàn)的cpg基序在胞嘧啶核苷酸上都是甲基化的。相比之下,抑制免疫應(yīng)答激活的核苷酸序列(命名為cpg中和,或cpg-n)在真核生物基因組中是過度出現(xiàn)的。最佳免疫刺激序列已發(fā)現(xiàn)是未甲基化的cpg二核苷酸,其側(cè)面為2個(gè)5'嘌呤和2個(gè)3'嘧啶。此外,在這個(gè)免疫刺激六聚體外的側(cè)面區(qū)域任選是富含鳥嘌呤的,以確保結(jié)合且攝取到靶細(xì)胞內(nèi)。
先天性免疫系統(tǒng)與適應(yīng)性免疫系統(tǒng)協(xié)同作用,以設(shè)定針對(duì)dna編碼蛋白質(zhì)的應(yīng)答。cpg-s序列誘導(dǎo)多克隆b細(xì)胞激活以及細(xì)胞因子表達(dá)和分泌的上調(diào)。受刺激的巨噬細(xì)胞分泌il-12、il-18、tnf-α、ifn-α、ifn-β和ifn-γ,受刺激的b細(xì)胞分泌il-6和一些il-12。在dna疫苗的質(zhì)粒骨架中cpg-s和cpg-n序列的操縱可以確保對(duì)于編碼抗原的免疫應(yīng)答的成功,并且驅(qū)動(dòng)針對(duì)th1表型的免疫應(yīng)答。如果病原體要求th應(yīng)答用于保護(hù),那么這是有用的。cpg-s序列也已用作外部佐劑,用于具有可變成功率的dna和重組蛋白質(zhì)疫苗接種。具有低甲基化的cpg基序的其他生物也已證實(shí)多克隆b細(xì)胞擴(kuò)增的刺激。然而,在這背后的機(jī)制可以比簡單甲基化更復(fù)雜–未發(fā)現(xiàn)低甲基化的鼠類dna設(shè)定免疫應(yīng)答。
dna的配制
dna免疫接種的效率可以通過針對(duì)降解穩(wěn)定dna且增加dna遞送到抗原呈遞細(xì)胞內(nèi)的效率得到改善。這可以通過用dna包被生物可降解的陽離子微粒(例如用十六烷基三甲基溴化銨配制的聚(丙交酯共乙交酯))來達(dá)到。此類dna包被微粒在產(chǎn)生ctl方面可以與重組痘苗病毒一樣有效,特別是當(dāng)與鋁混合時(shí)。直徑300nm的顆粒看起來對(duì)于通過抗原呈遞細(xì)胞攝取是最有效的。
施用
根據(jù)本發(fā)明的核酸構(gòu)建體和組合物可以以治療有效量施用于個(gè)體。有效量可以根據(jù)各種因素例如個(gè)體的狀況、重量、性別和年齡而改變。其他因素包括施用方式。
在一個(gè)實(shí)施方案中,根據(jù)本發(fā)明的核酸構(gòu)建體可以以dna、rna、lna、pna、ina、tina、hna、ana、cna、cena、gna、tna、缺口-聚體、混合-聚體(mix-mers)、嗎啉代類物質(zhì)或其任何組合的形式遞送給受試者。
在一個(gè)實(shí)施方案中,根據(jù)本發(fā)明的核酸構(gòu)建體可以以dna的形式遞送給受試者。
在另一個(gè)實(shí)施方案中,根據(jù)本發(fā)明的核酸構(gòu)建體可以以rna的形式遞送給受試者。因此,核酸構(gòu)建體可以在施用前轉(zhuǎn)錄成rna。
在另外一個(gè)實(shí)施方案中,根據(jù)本發(fā)明的核酸構(gòu)建體可以以蛋白質(zhì)的形式遞送給受試者。因此,核酸構(gòu)建體可以在施用前翻譯成蛋白質(zhì)。
在其中根據(jù)本發(fā)明的核酸構(gòu)建體以蛋白質(zhì)的形式遞送給受試者的實(shí)施方案中,蛋白質(zhì)可以已進(jìn)行修飾,以增加穩(wěn)定作用和/或最優(yōu)化進(jìn)入細(xì)胞內(nèi)的遞送。蛋白質(zhì)可以具有增加的穩(wěn)定性,原因在于二硫鍵的存在(例如us5,102,985用過氧化氫處理以還原形式的蛋白質(zhì)溶液,以90-96%得率生成具有分子內(nèi)二硫橋的蛋白質(zhì))、極性殘基中的增加、表面電荷最優(yōu)化、表面鹽橋、封裝(例如用中孔硅酸鹽),或蛋白質(zhì)可以與熱休克蛋白(例如hsp-60、hsp-70、hsp-90、hsp-20、hsp-27、hsp-84及其他)、hivtat易位結(jié)構(gòu)域、腺病毒纖維蛋白質(zhì)或任何其他蛋白質(zhì)或結(jié)構(gòu)域連接。
藥物或獸醫(yī)學(xué)組合物可以通過各種途徑提供給個(gè)體,例如皮下(sc或s.c.)、局部、經(jīng)口和肌內(nèi)(im或i.m.)。藥物組合物的施用經(jīng)口或腸胃外實(shí)現(xiàn)。腸胃外遞送的方法包括局部、動(dòng)脈內(nèi)(直接對(duì)于組織)、肌內(nèi)、大腦內(nèi)(ic或i.c.)、皮下、髓內(nèi)、鞘內(nèi)、心室內(nèi)、靜脈內(nèi)(iv或i.v.)、腹膜內(nèi)或鼻內(nèi)施用。本發(fā)明還具有提供用于在用組合物引發(fā)免疫應(yīng)答的方法中使用的合適局部、經(jīng)口、全身和腸胃外藥物制劑的目的。
例如,組合物可以在此類經(jīng)口劑型中施用,例如片劑、膠囊(各自包括限時(shí)釋放和持續(xù)釋放制劑)、丸劑、粉劑、顆粒劑、酏劑、酊劑、溶液、懸浮液、糖漿劑和乳狀劑,或通過注射。同樣地,它們還可以以靜脈內(nèi)(彈丸和輸注)、腹膜內(nèi)、皮下、局部連同或不連同阻塞、或肌內(nèi)形式施用,全部使用藥學(xué)領(lǐng)域普通技術(shù)人員眾所周知的形式。可以采用包括本文所述化合物中的任何的有效但無毒量的組合物。還包含其是本領(lǐng)域已知適合于配制可注射免疫原性肽組合物的任何和所有常規(guī)劑型,例如凍干形式和溶液、懸浮液或乳劑形式,需要時(shí),其包含常規(guī)藥學(xué)上可接受的載體、稀釋劑、防腐劑、佐劑、緩沖組分等。
在一個(gè)實(shí)施方案中,用于引發(fā)和/或后續(xù)加強(qiáng)疫苗的組合物作為緩慢或持續(xù)釋放制劑給予。
根據(jù)本發(fā)明的核酸構(gòu)建體或組合物的優(yōu)選施用方式包括但不限于全身施用,例如靜脈內(nèi)或皮下施用、真皮內(nèi)施用、肌內(nèi)施用、鼻內(nèi)施用、經(jīng)口施用、直腸施用、陰道施用、肺施用和一般地任何形式的粘膜施用。此外,在本發(fā)明中包括用于在本文中提及的任何施用形式的方法在本發(fā)明的范圍內(nèi)。
根據(jù)本發(fā)明的核酸構(gòu)建體或組合物可以施用一次,或任何次數(shù)例如2、3、4或5次。
在一個(gè)優(yōu)選實(shí)施方案中,核酸構(gòu)建體或組合物施用一次,隨后為合適疫苗的施用。
在另一個(gè)優(yōu)選實(shí)施方案中,核酸構(gòu)建體或組合物在施用疫苗前作為一系列施用進(jìn)行施用。此類系列可以包括每天、每第二天、每第三天、每第四天、每第五天、每第六天、每周、每兩周或每第三周施用核酸構(gòu)建體或組合物,總共1、2、3、4或5次。
在一個(gè)實(shí)施方案中,在首先施用核酸構(gòu)建體或組合物用于引發(fā)免疫應(yīng)答和其次施用疫苗用于加強(qiáng)之間的時(shí)間段是相隔至少1天、例如相隔至少2天、例如相隔3天、例如相隔至少4天、例如相隔5天、例如相隔至少6天、例如相隔7天、例如相隔至少8天、例如相隔9天、例如相隔至少10天、例如相隔15天、例如相隔至少20天、例如相隔25天。
用核酸構(gòu)建體或組合物引發(fā)因此預(yù)期通過施用疫苗得到進(jìn)一步加強(qiáng)。施用可以以不同于先前施用或類似于先前施用的形式或在不同于先前施用或類似于先前施用的身體部分中。
加強(qiáng)注射(shot)是同源或異源加強(qiáng)注射。同源加強(qiáng)注射是其中第一次和后續(xù)施用包括相同構(gòu)建體且更具體而言相同遞送媒介物。異源加強(qiáng)注射是其中等同構(gòu)建體包括在不同載體內(nèi)。
根據(jù)本發(fā)明的組合物的優(yōu)選施用形式是將組合物施用于身體區(qū)域,內(nèi)或外,最可能是給定感染的接受器(receptacle)。感染接受器是感染通過其接受的身體區(qū)域,例如關(guān)于流感,感染接受器是肺。
本發(fā)明的核酸構(gòu)建體或組合物可以施用于它對(duì)于其可以是有利的任何生物,特別是任何動(dòng)物例如脊椎動(dòng)物。組合物的施用裝置和方式適合于接受者落入本發(fā)明的范圍內(nèi)。
組合物的優(yōu)選接受者是哺乳動(dòng)物,并且在本發(fā)明的一個(gè)更優(yōu)選實(shí)施方案中,哺乳動(dòng)物選自:牛、豬、馬、綿羊、山羊、美洲駝、小鼠、大鼠、猴、犬、貓、雪貂和人。在最優(yōu)選的實(shí)施方案中,哺乳動(dòng)物是人。
組合物的另一個(gè)優(yōu)選接受者是來自鳥綱(鳥)的任何脊椎動(dòng)物,例如家雞(雞)。
本發(fā)明的一個(gè)實(shí)施方案包括進(jìn)一步包括第二種活性成分的組合物。第二種活性成分選自但不限于佐劑、抗生素、化學(xué)治療、抗變應(yīng)原、細(xì)胞因子、免疫系統(tǒng)的補(bǔ)體因子和共刺激分子。
本發(fā)明的另一個(gè)實(shí)施方案包括部分試劑盒,其中試劑盒包括根據(jù)上文中的任何的至少一種核酸構(gòu)建體或組合物,用于施用所述核酸構(gòu)建體或組合物的裝置,和關(guān)于如何做到這點(diǎn)的說明書。包括多個(gè)劑量的相同組合物或幾種不同組合物在本發(fā)明的范圍內(nèi)。在一個(gè)優(yōu)選實(shí)施方案中,部分試劑盒進(jìn)一步包括第二種活性成分。在一個(gè)更優(yōu)選的實(shí)施方案中,所述第二種活性成分是合適疫苗,即能夠加強(qiáng)通過所述免疫應(yīng)答的先前引發(fā)產(chǎn)生的免疫應(yīng)答的疫苗。
本發(fā)明進(jìn)一步包括用于加強(qiáng)動(dòng)物中的免疫應(yīng)答的方法,其包括給動(dòng)物施用根據(jù)上文中的任何的核酸構(gòu)建體或組合物,隨后施用合適疫苗,從而引發(fā)且加強(qiáng)受試者的免疫系統(tǒng)。
免疫應(yīng)答可以是但不限于下述應(yīng)答類型中的任何:mhc-i依賴性應(yīng)答、mhc-i和/或mhc-ii依賴性應(yīng)答、t細(xì)胞依賴性應(yīng)答、cd4+t細(xì)胞依賴性應(yīng)答、cd4+t細(xì)胞不依賴性應(yīng)答、cd8+t細(xì)胞依賴性應(yīng)答和b細(xì)胞依賴性免疫應(yīng)答。合適疫苗是在用根據(jù)本發(fā)明的核酸構(gòu)建體或組合物引發(fā)免疫系統(tǒng)后能夠加強(qiáng)免疫系統(tǒng)的那些。
在一個(gè)進(jìn)一步的實(shí)施方案中,本發(fā)明涉及治療有此需要的個(gè)體的方法,其包括施用如本文上文所述的組合物,以治療所述個(gè)體中的臨床狀況。
增加疫苗的效力
本發(fā)明的一個(gè)實(shí)施方案涉及編碼至少一種恒定鏈或其變體和至少一種抗原蛋白質(zhì)或肽或蛋白質(zhì)或肽的片段的核酸構(gòu)建體,其中至少一種抗原蛋白質(zhì)或肽或蛋白質(zhì)或肽的片段來自病毒、細(xì)菌或寄生蟲。
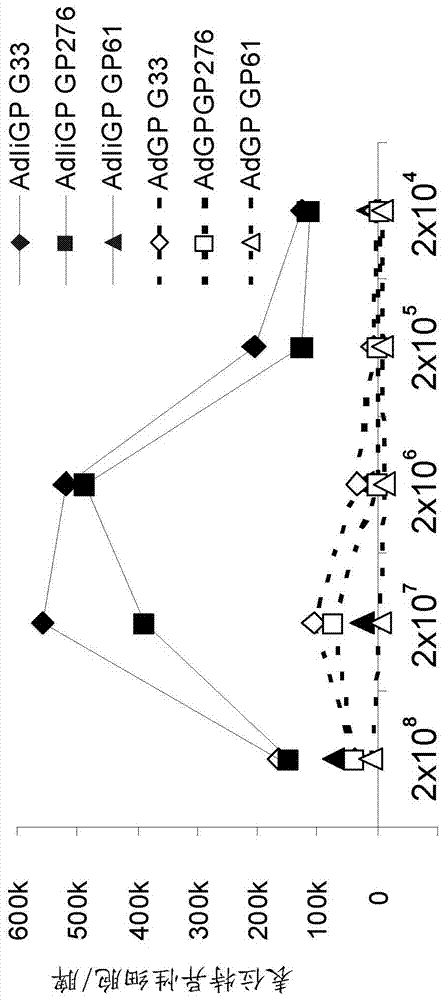
本文呈現(xiàn)的數(shù)據(jù)顯示使用包括恒定鏈或其變體的核酸構(gòu)建體開發(fā)引發(fā)-加強(qiáng)方案并不簡單。因此,如圖1中呈現(xiàn)的,包括恒定鏈和抗原的裸露dna構(gòu)建體(dna-iigp)能夠引發(fā)免疫應(yīng)答,而包括抗原但不含恒定鏈的裸露dna構(gòu)建體(dna-gp)不能引發(fā)免疫應(yīng)答。此外,圖12中呈現(xiàn)的數(shù)據(jù)顯示包括抗原的腺病毒載體(adgp)能夠引發(fā)特定(ad-iigp)但并非全部(ad-gp)免疫應(yīng)答,而圖13中呈現(xiàn)的數(shù)據(jù)顯示包括恒定鏈連同抗原的腺病毒載體(ad-iigp)在正常劑量和處理方案下不能引發(fā)任何(ad-iigp和ad-gp)免疫應(yīng)答。然而,可能的是通過使用較低劑量的ad-iigp用于引發(fā)來最優(yōu)化ad-iigp加強(qiáng)的ad-iigp引發(fā)(如圖14中所示)。
本發(fā)明的一個(gè)目的是提供編碼至少一種恒定鏈和病毒、細(xì)菌或寄生蟲抗原或其片段的核酸構(gòu)建體,其中所述恒定鏈?zhǔn)呛愣ㄦ湹淖凅w,用于引發(fā)免疫應(yīng)答,其中所述引發(fā)隨后為用癌癥疫苗的后續(xù)加強(qiáng)疫苗接種。所述恒定鏈的變體可以是如本文其他地方指定的任何變體,包括其中ii-keylrmk氨基酸殘基已通過例如缺失或置換加以改變,或其中clip區(qū)域的部分已通過例如缺失或置換加以改變的恒定鏈。
在一個(gè)實(shí)施方案中,本發(fā)明涉及核酸構(gòu)建體用于增加疫苗效力的用途。
在一個(gè)實(shí)施方案中,本發(fā)明公開了用于增加疫苗效力的方法,其包括步驟:
a.提供包括恒定鏈或其變體和抗原肽或其片段的核酸構(gòu)建體,
b.通過施用步驟a)的核酸構(gòu)建體引發(fā)受試者的免疫系統(tǒng),從而刺激所述受試者中的免疫應(yīng)答,和
c.通過施用合適疫苗加強(qiáng)步驟b)的免疫應(yīng)答。
在一個(gè)實(shí)施方案中,本發(fā)明公開了用于增加疫苗效力的方法,其包括步驟:
a.提供包括恒定鏈的變體和抗原肽或其片段的核酸構(gòu)建體,
b.通過施用步驟a)的核酸構(gòu)建體引發(fā)受試者的免疫系統(tǒng),從而刺激所述受試者中的免疫應(yīng)答,和
c.通過施用合適疫苗加強(qiáng)步驟b)的免疫應(yīng)答,
其中所述恒定鏈的變體包括通過例如缺失或置換的ii-keylrmk氨基酸殘基的改變。
在一個(gè)實(shí)施方案中,本發(fā)明公開了用于增加疫苗效力的方法,其包括步驟:
a.提供包括恒定鏈的變體和抗原肽或其片段的核酸構(gòu)建體,
b.通過施用步驟a)的核酸構(gòu)建體引發(fā)受試者的免疫系統(tǒng),從而刺激所述受試者中的免疫應(yīng)答,和
c.通過施用合適疫苗加強(qiáng)步驟b)的免疫應(yīng)答,
其中所述恒定鏈的變體包括通過例如缺失或置換的clip區(qū)域改變。
在一個(gè)實(shí)施方案中,本發(fā)明公開了用于增加疫苗效力的方法,其包括步驟:
a.提供包括恒定鏈的變體和抗原肽或其片段的核酸構(gòu)建體,
b.通過施用步驟a)的核酸構(gòu)建體引發(fā)受試者的免疫系統(tǒng),從而刺激所述受試者中的免疫應(yīng)答,和
c.通過施用合適疫苗加強(qiáng)步驟b)的免疫應(yīng)答,
其中所述恒定鏈的變體不包括前17個(gè)氨基酸。
在另一個(gè)實(shí)施方案中,本發(fā)明涉及核酸構(gòu)建體用于引發(fā)免疫應(yīng)答的用途。
在一個(gè)實(shí)施方案中,本發(fā)明公開了用于引發(fā)免疫應(yīng)答的方法,其包括步驟:
a.提供包括恒定鏈或其變體和抗原肽或其片段的核酸構(gòu)建體,
b.通過施用步驟a)的核酸構(gòu)建體引發(fā)受試者的免疫系統(tǒng),從而刺激所述受試者中的免疫應(yīng)答,和
c.通過施用合適疫苗加強(qiáng)步驟b)的免疫應(yīng)答。
在一個(gè)實(shí)施方案中,本發(fā)明公開了用于引發(fā)免疫應(yīng)答的方法,其包括步驟:
a.提供包括恒定鏈的變體和抗原肽或其片段的核酸構(gòu)建體,
b.通過施用步驟a)的核酸構(gòu)建體引發(fā)受試者的免疫系統(tǒng),從而刺激所述受試者中的免疫應(yīng)答,和
c.通過施用合適疫苗加強(qiáng)步驟b)的免疫應(yīng)答,
其中所述恒定鏈的變體包括通過例如缺失或置換的ii-keylrmk氨基酸殘基改變。
在一個(gè)實(shí)施方案中,本發(fā)明公開了用于引發(fā)免疫應(yīng)答的方法,其包括步驟:
a.提供包括恒定鏈的變體和抗原肽或其片段的核酸構(gòu)建體,
b.通過施用步驟a)的核酸構(gòu)建體引發(fā)受試者的免疫系統(tǒng),從而刺激所述受試者中的免疫應(yīng)答,和
c.通過施用合適疫苗加強(qiáng)步驟b)的免疫應(yīng)答,
其中所述恒定鏈的變體包括通過例如缺失或置換的clip區(qū)域改變。
在一個(gè)實(shí)施方案中,本發(fā)明公開了用于引發(fā)免疫應(yīng)答的方法,其包括步驟:
a.提供包括恒定鏈的變體和抗原肽或其片段的核酸構(gòu)建體,
b.通過施用步驟a)的核酸構(gòu)建體引發(fā)受試者的免疫系統(tǒng),從而刺激所述受試者中的免疫應(yīng)答,和
c.通過施用合適疫苗加強(qiáng)步驟b)的免疫應(yīng)答,
其中所述恒定鏈的變體不包括前17個(gè)氨基酸。
增加癌癥疫苗的效力
本發(fā)明的一個(gè)實(shí)施方案涉及編碼至少一種恒定鏈或其變體和至少一種抗原蛋白質(zhì)或肽或蛋白質(zhì)或肽的片段的核酸構(gòu)建體,其中至少一種抗原蛋白質(zhì)或肽或蛋白質(zhì)或肽的片段來自癌癥特異性多肽或癌抗原。
本發(fā)明的一個(gè)目的是提供編碼至少一種恒定鏈和癌抗原或其片段的核酸構(gòu)建體,其中所述恒定鏈以其天然、野生型形式,用于引發(fā)免疫應(yīng)答,其中所述引發(fā)隨后為用癌癥疫苗的后續(xù)加強(qiáng)疫苗接種。
本發(fā)明的另外一個(gè)目的是提供編碼至少一種恒定鏈和癌抗原或其片段的核酸構(gòu)建體,其中所述恒定鏈?zhǔn)呛愣ㄦ湹淖凅w,用于引發(fā)免疫應(yīng)答,其中所述引發(fā)隨后為用癌癥疫苗的后續(xù)加強(qiáng)疫苗接種。所述恒定鏈的變體可以是如本文其他地方指定的任何變體,包括其中ii-keylrmk氨基酸殘基已通過例如缺失或置換加以改變,或其中clip區(qū)域的部分已通過例如缺失或置換加以改變,或其中前17個(gè)氨基酸已缺失的恒定鏈。
由此可見當(dāng)用于加強(qiáng)免疫應(yīng)答的隨后施用的疫苗是癌癥疫苗時(shí),由根據(jù)本發(fā)明的核酸構(gòu)建體編碼的恒定鏈可以以其天然、野生型形式,或它可以是恒定鏈的變體。
在一個(gè)實(shí)施方案中,本發(fā)明涉及核酸構(gòu)建體用于增加癌癥疫苗效力的用途。
在一個(gè)實(shí)施方案中,本發(fā)明公開了用于增加癌癥疫苗效力的方法,其包括步驟:
a.提供包括恒定鏈或其變體和癌癥特異性抗原肽或其片段的核酸構(gòu)建體,
b.通過施用步驟a)的核酸構(gòu)建體引發(fā)受試者的免疫系統(tǒng),從而刺激所述受試者中的免疫應(yīng)答,和
c.通過施用合適的癌癥疫苗加強(qiáng)步驟b)的免疫應(yīng)答。
在一個(gè)實(shí)施方案中,本發(fā)明公開了用于增加癌癥疫苗效力的方法,其包括步驟:
a.提供包括恒定鏈和癌癥特異性抗原肽或其片段的核酸構(gòu)建體,
b.通過施用步驟a)的核酸構(gòu)建體引發(fā)受試者的免疫系統(tǒng),從而刺激所述受試者中的免疫應(yīng)答,和
c.通過施用合適的癌癥疫苗加強(qiáng)步驟b)的免疫應(yīng)答,
其中所述恒定鏈以其天然、野生型形式。
在一個(gè)實(shí)施方案中,本發(fā)明公開了用于增加癌癥疫苗效力的方法,其包括步驟:
a.提供包括恒定鏈的變體和癌癥特異性抗原肽或其片段的核酸構(gòu)建體,
b.通過施用步驟a)的核酸構(gòu)建體引發(fā)受試者的免疫系統(tǒng),從而刺激所述受試者中的免疫應(yīng)答,和
c.通過施用合適的癌癥疫苗加強(qiáng)步驟b)的免疫應(yīng)答,
其中所述恒定鏈的變體包括通過例如缺失或置換的ii-keylrmk氨基酸殘基的改變,和/或通過例如缺失或置換的clip區(qū)域部分的改變,和/或ii的前17個(gè)氨基酸的缺失。
在另一個(gè)實(shí)施方案中,本發(fā)明涉及核酸構(gòu)建體用于引發(fā)免疫應(yīng)答的用途。
在一個(gè)實(shí)施方案中,本發(fā)明公開了用于引發(fā)免疫應(yīng)答的方法,其包括步驟:
a.提供包括恒定鏈或其變體和癌癥特異性抗原肽或其片段的核酸構(gòu)建體,
b.通過施用步驟a)的核酸構(gòu)建體引發(fā)受試者的免疫系統(tǒng),從而刺激所述受試者中的免疫應(yīng)答,和
c.通過施用合適的癌癥疫苗加強(qiáng)步驟b)的免疫應(yīng)答。
在一個(gè)實(shí)施方案中,本發(fā)明公開了用于引發(fā)免疫應(yīng)答的方法,其包括步驟:
a.提供包括恒定鏈和癌癥特異性抗原肽或其片段的核酸構(gòu)建體,
b.通過施用步驟a)的核酸構(gòu)建體引發(fā)受試者的免疫系統(tǒng),從而刺激所述受試者中的免疫應(yīng)答,和
c.通過施用合適的癌癥疫苗加強(qiáng)步驟b)的免疫應(yīng)答,
其中所述恒定鏈以其天然、野生型形式。
在一個(gè)實(shí)施方案中,本發(fā)明公開了用于引發(fā)免疫應(yīng)答的方法,其包括步驟:
a.提供包括恒定鏈的變體和癌癥特異性抗原肽或其片段的核酸構(gòu)建體,
b.通過施用步驟a)的核酸構(gòu)建體引發(fā)受試者的免疫系統(tǒng),從而刺激所述受試者中的免疫應(yīng)答,和
c.通過施用合適的癌癥疫苗加強(qiáng)步驟b)的免疫應(yīng)答,
其中所述恒定鏈的變體包括通過例如缺失或置換的ii-keylrmk氨基酸殘基的改變,和/或通過例如缺失或置換的clip區(qū)域部分的改變,和/或ii的前17個(gè)氨基酸的缺失。
增加針對(duì)異常生理學(xué)應(yīng)答的疫苗的效力
本發(fā)明的一個(gè)目的是提供編碼至少一種恒定鏈和與異常生理學(xué)應(yīng)答相關(guān)的多肽或其片段的核酸構(gòu)建體,其中所述恒定鏈以其天然、野生型形式,用于引發(fā)免疫應(yīng)答,其中所述引發(fā)隨后為用針對(duì)所述異常生理學(xué)應(yīng)答的疫苗的后續(xù)加強(qiáng)疫苗接種。
本發(fā)明的另外一個(gè)目的是提供編碼至少一種恒定鏈和與異常生理學(xué)應(yīng)答相關(guān)的多肽或其片段的核酸構(gòu)建體,其中所述恒定鏈?zhǔn)呛愣ㄦ湹淖凅w,用于引發(fā)免疫應(yīng)答,其中所述引發(fā)隨后為用針對(duì)所述異常生理學(xué)應(yīng)答的疫苗的后續(xù)加強(qiáng)疫苗接種。所述恒定鏈的變體可以是如本文其他地方指定的任何變體,包括其中ii-keylrmk氨基酸殘基已通過例如缺失或置換加以改變,或其中clip區(qū)域的部分已通過例如缺失或置換加以改變,或其中前17個(gè)氨基酸已缺失的恒定鏈。
由此可見當(dāng)用于加強(qiáng)免疫應(yīng)答的隨后施用的疫苗是針對(duì)所述異常生理學(xué)應(yīng)答的疫苗時(shí),由根據(jù)本發(fā)明的核酸構(gòu)建體編碼的恒定鏈可以以其天然、野生型形式,或它可以是恒定鏈的變體。
在一個(gè)實(shí)施方案中,本發(fā)明涉及核酸構(gòu)建體用于增加針對(duì)異常生理學(xué)應(yīng)答的疫苗效力的用途。
在另一個(gè)實(shí)施方案中,本發(fā)明涉及核酸構(gòu)建體用于引發(fā)免疫應(yīng)答的用途。
在一個(gè)實(shí)施方案中,本發(fā)明公開了用于增加針對(duì)異常生理學(xué)應(yīng)答的疫苗效力的方法,其包括步驟:
a.提供包括恒定鏈或其變體和與異常生理學(xué)應(yīng)答相關(guān)的抗原肽或其片段的核酸構(gòu)建體,
b.通過施用步驟a)的核酸構(gòu)建體引發(fā)受試者的免疫系統(tǒng),從而刺激所述受試者中的免疫應(yīng)答,和
c.通過施用針對(duì)異常生理學(xué)應(yīng)答的合適疫苗加強(qiáng)步驟b)的免疫應(yīng)答。
在一個(gè)實(shí)施方案中,本發(fā)明公開了用于增加針對(duì)異常生理學(xué)應(yīng)答的疫苗效力的方法,其包括步驟:
a.提供包括恒定鏈和與異常生理學(xué)應(yīng)答相關(guān)的抗原肽或其片段的核酸構(gòu)建體,
b.通過施用步驟a)的核酸構(gòu)建體引發(fā)受試者的免疫系統(tǒng),從而刺激所述受試者中的免疫應(yīng)答,和
c.通過施用針對(duì)異常生理學(xué)應(yīng)答的合適疫苗加強(qiáng)步驟b)的免疫應(yīng)答,
其中所述恒定鏈以其天然、野生型形式。
在一個(gè)實(shí)施方案中,本發(fā)明公開了用于增加針對(duì)異常生理學(xué)應(yīng)答的疫苗效力的方法,其包括步驟:
a.提供包括恒定鏈的變體和與異常生理學(xué)應(yīng)答相關(guān)的抗原肽或其片段的核酸構(gòu)建體,
b.通過施用步驟a)的核酸構(gòu)建體引發(fā)受試者的免疫系統(tǒng),從而刺激所述受試者中的免疫應(yīng)答,和
c.通過施用針對(duì)異常生理學(xué)應(yīng)答的合適疫苗加強(qiáng)步驟b)的免疫應(yīng)答,
其中所述恒定鏈的變體包括通過例如缺失或置換的ii-keylrmk氨基酸殘基的改變,和/或通過例如缺失或置換的clip區(qū)域部分的改變,和/或ii的前17個(gè)氨基酸的缺失。
疫苗類型
本發(fā)明的一個(gè)方面涉及受試者中的免疫應(yīng)答引發(fā),通過施用包括ii連接的抗原的核酸構(gòu)建體來進(jìn)行,隨后為通過給相同受試者施用合適疫苗達(dá)到的后續(xù)加強(qiáng)。
根據(jù)本發(fā)明的合適疫苗具有與用于引發(fā)免疫應(yīng)答的核酸構(gòu)建體共同的至少一個(gè)等同特征。所述等同特征可以包括在恒定鏈的部分或全部、抗原肽的部分或全部、主鏈結(jié)構(gòu)的部分或全部例如啟動(dòng)子區(qū)的部分或全部、增強(qiáng)子的部分或全部、終止子的部分或全部、多聚腺苷酸尾的部分或全部、接頭的部分或全部、多聚接頭的部分或全部、操作接頭的部分或全部、多克隆位點(diǎn)(mcs)的部分或全部、標(biāo)記的部分或全部、終止密碼子的部分或全部、內(nèi)部核糖體進(jìn)入位點(diǎn)(ires)的部分或全部和用于整合的宿主同源序列或其他限定元件的部分或全部中。
在一個(gè)優(yōu)選實(shí)施方案中,等同特征是抗原肽或獨(dú)特輔助t細(xì)胞表位的部分或全部。在一個(gè)最優(yōu)選的實(shí)施方案中,等同特征是抗原肽的部分或全部。
在另一個(gè)優(yōu)選實(shí)施方案中,等同特征是恒定鏈的部分或全部。
疫苗可以被視為常規(guī)或創(chuàng)新的。本文引用的疫苗類型中的任何可以在根據(jù)本發(fā)明的后續(xù)加強(qiáng)步驟中使用。
常規(guī)疫苗或第一代疫苗依賴于完整生物;已被殺死的致病菌株,或具有減毒致病性的菌株。
分子生物學(xué)技術(shù)已用于開發(fā)新疫苗,基于來自致病生物的個(gè)別抗原蛋白質(zhì)的第二代疫苗。概念上,抗原肽而不是完整生物的使用將避免致病性,同時(shí)提供包含最具免疫原性的抗原的疫苗。這些包括基于眾所周知的滅活毒性化合物的基于類毒素的疫苗,和基于滅活或減毒致病菌株的片段的亞單位疫苗。
綴合物疫苗:特定細(xì)菌具有弱免疫原性的多糖外衣殼。通過使這些外衣殼與蛋白質(zhì)(例如毒素)連接,免疫系統(tǒng)可以導(dǎo)致識(shí)別多糖,如同它是蛋白質(zhì)抗原。
重組載體疫苗:通過使一種微生物的生理學(xué)和另一種的dna組合,免疫可以針對(duì)具有復(fù)雜感染過程的疾病產(chǎn)生。
合成疫苗主要或整個(gè)由合成肽、碳水化合物或抗原組成。
dna(或遺傳)疫苗或第三代疫苗是用于開發(fā)預(yù)防和治療疫苗的新的和有希望的候選物。dna疫苗由dna的小環(huán)狀段(質(zhì)粒)構(gòu)成,其已進(jìn)行遺傳改造,以產(chǎn)生來自微生物的一種或多種抗原。將疫苗dna注射到機(jī)體的細(xì)胞內(nèi),其中宿主細(xì)胞的“內(nèi)部機(jī)制”“閱讀”dna,并且將其轉(zhuǎn)變成致病蛋白質(zhì)。因?yàn)檫@些蛋白質(zhì)被識(shí)別為外來的,所以它們通過宿主細(xì)胞加工且在其表面上展示,以改變免疫系統(tǒng),這隨后觸發(fā)一系列免疫應(yīng)答。確保免疫應(yīng)答的強(qiáng)度通過載體(即裸露dna、病毒載體、活減毒病毒等)效力、抗原表達(dá)水平和重組抗原自身(即高或低親和力mhc結(jié)合物、對(duì)于或多或少有限的t或b細(xì)胞儲(chǔ)庫選擇的結(jié)構(gòu)決定簇等)的組合決定。免疫記憶的有效誘導(dǎo)要求或獲益于cd4+(輔助細(xì)胞)t細(xì)胞與介導(dǎo)免疫記憶的許多效應(yīng)的cd8+(細(xì)胞毒性)t細(xì)胞和b細(xì)胞的相互作用,這一般是真實(shí)的。
在本發(fā)明的一個(gè)實(shí)施方案中,用根據(jù)本發(fā)明的核酸構(gòu)建體引發(fā)免疫應(yīng)答隨后為用于加強(qiáng)所述免疫應(yīng)答的第一代或常規(guī)疫苗的后續(xù)施用。
在本發(fā)明的一個(gè)實(shí)施方案中,用根據(jù)本發(fā)明的核酸構(gòu)建體引發(fā)免疫應(yīng)答隨后為用于加強(qiáng)所述免疫應(yīng)答的第二代疫苗的后續(xù)施用。
在本發(fā)明的一個(gè)實(shí)施方案中,用根據(jù)本發(fā)明的核酸構(gòu)建體引發(fā)免疫應(yīng)答隨后為用于加強(qiáng)所述免疫應(yīng)答的第三代或dna疫苗的后續(xù)施用。
在dna疫苗構(gòu)建體中使用恒定鏈以增加免疫原性是本領(lǐng)域眾所周知的。在本發(fā)明的一個(gè)實(shí)施方案中,用根據(jù)本發(fā)明的核酸構(gòu)建體引發(fā)免疫應(yīng)答隨后為用于加強(qiáng)所述免疫應(yīng)答的dna疫苗的后續(xù)施用,所述dna疫苗包括恒定鏈或其變體。
在本發(fā)明的一個(gè)實(shí)施方案中,用根據(jù)本發(fā)明的核酸構(gòu)建體引發(fā)免疫應(yīng)答隨后為用于加強(qiáng)所述免疫應(yīng)答的腺病毒疫苗的后續(xù)施用。
疫苗可以進(jìn)一步是單價(jià)的(monovalent)(也稱為一價(jià)的(univalent))或多價(jià)的(multivalent)(也稱為多價(jià)的(polyvalent))。單價(jià)疫苗設(shè)計(jì)為針對(duì)單一抗原或單一微生物免疫接種。多價(jià)或多價(jià)體疫苗設(shè)計(jì)為針對(duì)相同微生物的2種或更多種菌株,或針對(duì)2種或更多種微生物免疫接種。
附圖詳述
圖1:用基于ii鏈的裸露dna疫苗的dna-引發(fā)顯著增強(qiáng)在用高度有效的病毒載體的后續(xù)加強(qiáng)后的病毒特異性cd8+t細(xì)胞生成。小鼠用dna-iigp、dna-gp進(jìn)行基因槍免疫接種2次,相隔3周,或不予以處理。在最后一次免疫接種后3周,所有小鼠在右后足墊中用2x107ifuad5-iigp注射,并且4周后,處死動(dòng)物,并且如圖1中所述分析脾細(xì)胞。表位特異性ifn-γ+cd8+t細(xì)胞數(shù)目作為平均值±se(n=5只小鼠/組)呈現(xiàn)。*指示相對(duì)于僅用ad5-iigp疫苗接種的小鼠的統(tǒng)計(jì)顯著性(曼-懷二氏秩和檢驗(yàn))。描述了來自2次相似實(shí)驗(yàn)之一的結(jié)果。
圖2:ii序列中的結(jié)構(gòu)域和測試突變定位。wtii中的結(jié)構(gòu)域在條上描述。ess;胞內(nèi)體分選信號(hào),tm;跨膜結(jié)構(gòu)域,key;肽呈遞增強(qiáng)區(qū)域,clip;ii類結(jié)合的恒定鏈肽,trim:三聚結(jié)構(gòu)域。ii中的缺失突變和置換程度在條下標(biāo)記。a;ad-δ17iigp,b;ad-iiltmgp,c;ad-iiutmgp,d;ad-δ50iigp,e;ad-ii1-201gp,f;ad-ii1-118gp,g;ad-ii1-105gp,h;ad-iiclipgp,i;ad-iikeygp,j;ad-ii51-118gp。
圖3:ii顯著增加siinfekl/h-2kbova衍生表位的細(xì)胞表面呈遞。骨髓衍生的樹突細(xì)胞用ad-ova、ad-iiova或ad-iigp(陰性對(duì)照)轉(zhuǎn)染,并且表面對(duì)于mhcii類(染色成熟樹突細(xì)胞)和用siinfekl/h-2kb特異性抗體(ova表位)染色。
圖4:ii僅以順式工作。a)來自ad-iigp和ad-ii+gp載體的ii的表達(dá);在用50moiad-iigp和ad-ii+gp感染的cos7細(xì)胞中將ii表達(dá)對(duì)gapdh進(jìn)行標(biāo)準(zhǔn)化。b)tcr318gp33限制響應(yīng)ad-gp、ad-iigp或ad-ii+gp轉(zhuǎn)導(dǎo)的bmdc(骨髓衍生的樹突細(xì)胞)的t細(xì)胞增殖。
圖5:n末端缺失和置換不影響ii刺激能力。tcr318gp33限制響應(yīng)ad-gp、ad-iigp、ad-δ17iigp、ad-iiltmgp、ad-iiutmgp、ad-δ50iigp轉(zhuǎn)導(dǎo)的bmdc(骨髓衍生的樹突細(xì)胞)的t細(xì)胞增殖。
圖6:c末端缺失和置換不影響ii刺激能力。tcr318gp33限制響應(yīng)ad-gp、ad-iigp、ad-ii1-205gp、adii1-118gp和ad-ii1-105gp轉(zhuǎn)導(dǎo)的bmdc(骨髓衍生的樹突細(xì)胞培養(yǎng)系統(tǒng))的t細(xì)胞增殖。
圖7:僅n和c末端缺失減少ii刺激能力。tcr318gp33限制響應(yīng)ad-gp、ad-iigp、ad-iiclipgp、ad-iikeygp和ad-ii51-118gp轉(zhuǎn)導(dǎo)的bmdc(骨髓衍生的樹突細(xì)胞培養(yǎng)系統(tǒng))的t細(xì)胞增殖。
圖8:ad-iigp和ad-gp疫苗的劑量應(yīng)答。小鼠組用在指示菌株中的指示疫苗進(jìn)行疫苗接種。在疫苗接種后14天,處死小鼠,并且用指示表位刺激脾細(xì)胞。通過細(xì)胞內(nèi)染色和facs分析測定特異性cd8+脾細(xì)胞的總數(shù)目。數(shù)據(jù)顯示ad-iigp在極低劑量下誘導(dǎo)應(yīng)答,并且因此用低劑量ad-iigp(或任何抗原)引發(fā)和用更高劑量ad-iigp(或任何抗原)后續(xù)加強(qiáng)可以應(yīng)用于同源引發(fā)-加強(qiáng)方案。
圖9:ad-gp、ad-iigp和ad-iiclipgp用于mhcii類呈遞(cd4+t細(xì)胞的刺激)的比較。smartagp61-80限制響應(yīng)ad-gp、ad-iigp和ad-iiclipgp轉(zhuǎn)導(dǎo)的bmdc的t細(xì)胞呈遞顯示增加的ad-iiclipgp的mhcii抗原呈遞。
圖10:在體內(nèi)時(shí)間-過程研究中的ad-gp、ad-iigp、ad-gplamp-1和ad-iiδ17gp比較。
圖11:ad-gp、ad-iigp、ad-iiδ17gp、ad-iikeygp、ad-iiclipgp、ad-ii1-117gp和ad-ii1-199gp體內(nèi)應(yīng)答的比較。
圖12:ad-gp能夠引發(fā)后續(xù)ad-iigp加強(qiáng)。3組c57bl/6小鼠用ad-gp進(jìn)行疫苗接種。60天后,這些小鼠不受干擾,用ad-gp進(jìn)行疫苗接種,或用ad-iigp進(jìn)行疫苗接種。包括用ad-gp進(jìn)行疫苗接種的第4組小鼠。第一次疫苗接種后120天,處死小鼠,并且通過用所述肽的先體外后體內(nèi)再刺激和用于干擾素-γ產(chǎn)生的細(xì)胞內(nèi)染色來定量識(shí)別指示表位的抗原特異性細(xì)胞。
圖13:ad-iigp不能引發(fā)后續(xù)ad-gp或ad-iigp加強(qiáng)。3組c57bl/6小鼠用ad-iigp進(jìn)行疫苗接種。60天后,這些小鼠不受干擾,用ad-gp進(jìn)行疫苗接種,或用ad-iigp進(jìn)行疫苗接種。包括用ad-iigp進(jìn)行疫苗接種的第4組小鼠。第一次疫苗接種后120天,處死小鼠,并且通過用所述肽的先體外后體內(nèi)再刺激和用于干擾素-γ產(chǎn)生的細(xì)胞內(nèi)染色來定量識(shí)別指示表位的抗原特異性細(xì)胞。這顯示ad-iigp引發(fā)不能用ad-iigp加強(qiáng),而dna-iigp引發(fā)可以用ad-iigp加強(qiáng)(參見圖1)。
圖14:ad-gp和adii-gp疫苗的劑量應(yīng)答。小鼠組用在指示菌株中的指示疫苗進(jìn)行疫苗接種。在疫苗接種后14天,處死小鼠,并且用指示表位刺激脾細(xì)胞。通過細(xì)胞內(nèi)染色和facs分析測定cd8+脾細(xì)胞的總數(shù)目。
圖15:與包括殘基50–215(ii50-215)的恒定鏈的變體偶聯(lián)的甘露糖受體進(jìn)一步與腺病毒纖維蛋白質(zhì)偶聯(lián)。腺病毒纖維蛋白質(zhì)(ad纖維)可以源自腺病毒的任何血清型。甘露糖受體可以是來自甘露糖受體的一個(gè)或多個(gè)結(jié)構(gòu)域。ii可以是ii的變體或全長。ag=抗原。
本發(fā)明的實(shí)施例
本發(fā)明現(xiàn)在將參考下述實(shí)施例進(jìn)一步舉例說明。應(yīng)當(dāng)理解下述僅作為例子并且可以做出細(xì)節(jié)中的修飾,同時(shí)仍落入本發(fā)明的范圍內(nèi)。
實(shí)施例1:用基于ii鏈的裸露dna疫苗引發(fā)顯著加強(qiáng)在用最優(yōu)化病毒載體的后續(xù)加強(qiáng)后的病毒特異性cd8+t細(xì)胞生成。
用裸露dna疫苗(即核酸構(gòu)建體)引發(fā)顯示加強(qiáng)通過用ad5(腺病毒血清型5)載體的后續(xù)免疫接種產(chǎn)生的免疫應(yīng)答。用dna-iigp(表達(dá)與恒定鏈(ii)融合的lcmv(淋巴細(xì)胞性脈絡(luò)叢腦膜炎病毒)糖蛋白(gp)的dna構(gòu)建體)引發(fā)在本文中證實(shí)顯著增強(qiáng)通過在腺病毒血清型5載體(ad5-iigp)中遞送的相同基因構(gòu)建體誘導(dǎo)的cd8+t細(xì)胞應(yīng)答,提供關(guān)于在未來異源免疫接種(“引發(fā)-加強(qiáng)”)方案中包括基于ii鏈的dna構(gòu)建體的強(qiáng)論據(jù)。
我們的研究顯示ii鏈連接的免疫增強(qiáng)效應(yīng)并不限于ad5載體,還與dna平臺(tái)相關(guān)。此外,考慮ii鏈增強(qiáng)超過一種表位的呈遞的事實(shí),這提出基于ii鏈的dna疫苗作為用于各種異源引發(fā)-加強(qiáng)方案的非常有希望的候選物。
結(jié)果和討論
改善誘導(dǎo)的t細(xì)胞記憶的一種方法是通過異源引發(fā)-加強(qiáng)方案,例如裸露dna引發(fā)隨后為載體加強(qiáng)。因此具有在我們實(shí)驗(yàn)室中的合適載體——表達(dá)與p31ii鏈融合的lcmvgp的復(fù)制缺陷型腺病毒(ad5-iigp),在實(shí)驗(yàn)上測試了這種可能性。首先,我們執(zhí)行了標(biāo)準(zhǔn)dna疫苗接種,用dna-iigp或dna-gp的基因槍疫苗接種2次,相隔3周。第二次dna疫苗接種后3周,通過在右后足墊中接種2x107ifuad5-iigp免疫接種小鼠組和匹配對(duì)照,并且4周后,通過關(guān)于ifn-γ的iccs和流式細(xì)胞術(shù)計(jì)數(shù)脾中的病毒特異性cd8+t細(xì)胞數(shù)目。用融合的dna構(gòu)建體引發(fā)的小鼠包含比未引發(fā)小鼠明顯更多的gp33-41和gp276-286特異性ifn-γ+cd8+t細(xì)胞,并且對(duì)于gp92-101特異性細(xì)胞注意到相似趨勢,盡管在這種情況下差異不是統(tǒng)計(jì)上顯著的。相比之下,在不存在ii的情況下用編碼gp的裸露dna引發(fā)對(duì)通過用ad5-iigp的后續(xù)免疫接種誘導(dǎo)的gp特異性記憶cd8+t細(xì)胞水平具有很少作用(圖1)。應(yīng)當(dāng)指出觀察到的誘導(dǎo)ii作用不反映疫苗接種小鼠的免疫反應(yīng)性的非特異性加強(qiáng),因?yàn)橛脙H包括ii但不含gp的載體的dna引發(fā)對(duì)隨后用腺病毒載體接種的小鼠中的gp特異性cd8+t細(xì)胞水平?jīng)]有作用(數(shù)據(jù)未顯示)。
我們已顯示使用改良dna載體作為異源引發(fā)-加強(qiáng)方案的一部分將顯著加強(qiáng)通過已最優(yōu)化的病毒載體誘導(dǎo)的應(yīng)答(holst等人,2008)。這強(qiáng)烈指出甚至基于非常免疫原性載體的免疫接種也可以通過用基于ii鏈的裸露dna疫苗最初引發(fā)宿主得到進(jìn)一步改善。總之,因?yàn)榕c抗原的ii鏈融合物將導(dǎo)致用于廣泛cd8+t細(xì)胞應(yīng)答的引發(fā),所以基于ii鏈的dna疫苗應(yīng)代表關(guān)于針對(duì)快速突變病毒的預(yù)防策略作為異源引發(fā)-加強(qiáng)方案的一部分的明確優(yōu)點(diǎn)。
材料與方法
小鼠。c57bl/6(b6)野生型小鼠得自taconicm&b(ry,丹麥)。穿孔蛋白缺陷b6小鼠由最初得自thejacksonlaboratory(barharbor,me)的育種對(duì)當(dāng)?shù)嘏嘤?–10周大的小鼠在所有實(shí)驗(yàn)中使用,并且來自外部來源的動(dòng)物在使用前總是允許適應(yīng)當(dāng)?shù)丨h(huán)境至少1周。所有動(dòng)物在無特殊病原體的條件下飼養(yǎng),如通過標(biāo)記(sentinels)篩選驗(yàn)證的。所有動(dòng)物實(shí)驗(yàn)根據(jù)國家指導(dǎo)進(jìn)行。
dna疫苗構(gòu)建和免疫接種程序。dna疫苗使用真核生物表達(dá)載體paccmv.plpa產(chǎn)生,所述paccmv.plpa包含鼠類恒定鏈隨后為lcmv的gp或單獨(dú)的lcmvgp。構(gòu)建體如近期描述的生成(holst等人,2008)。大腸桿菌菌株xl1-blue(stratagene,usa)通過電穿孔用構(gòu)建體進(jìn)行轉(zhuǎn)化。使用循環(huán)測序、bigdyeterminator和abi310遺傳分析儀(abiprism,usa)的dna測序鑒定陽性克隆。引物得自tag,copenhagen,丹麥。使用qiagenmaxiprep(qiagen,usa)產(chǎn)生大規(guī)模dna制劑。
基因槍免疫接種。將dna以2μgdna/mg金的濃度包被到1.6nm金顆粒上,并且將dna/金復(fù)合物包被到塑料管上,從而使得0.5mg金遞送至小鼠/注射(1μgdna/注射)。這些程序根據(jù)制造商的說明書執(zhí)行(biorad,ca,usa)(bartholdy等人,2003)。小鼠使用采用壓縮氦(400psi)作為粒子原運(yùn)力的手提式基因槍裝置在腹部皮膚上進(jìn)行免疫接種。除非另有說明,否則小鼠用3-4周的間隔免疫接種2次,并且隨后在進(jìn)一步攻擊/研究前允許休息3周。
病毒。lcmv的阿姆斯特朗(armstrong)毒株克隆13在大多數(shù)實(shí)驗(yàn)中使用。除非另有說明,否則待感染的小鼠接受在0.3ml的i.v.注射中的105pfu,或在右后足墊(f.p.)中在0.03ml中的20pfu克隆13劑量。對(duì)于i.c.注射,小鼠接受在0.03ml的體積中的20pfu的向神經(jīng)阿姆斯特朗克隆53b。如近期描述的(holst等人,2008)產(chǎn)生且滴定編碼恒定鏈連接的gp的復(fù)制缺陷型腺病毒(ad5-iigp)。
病毒滴定。通過如先前描述的(battegay等人,1991)免疫焦點(diǎn)測定來測定器官病毒滴度。
cd4+和cd8+t細(xì)胞的體內(nèi)耗盡。使用抗cd4(克隆gk1.5)和抗cd8mabs(克隆53.6.72)。在相對(duì)于感染的第-1和0天時(shí),待耗盡細(xì)胞的小鼠腹膜內(nèi)接受在0.3mlpbs的體積中的200□g劑量;而對(duì)于假處理,使用純化的大鼠igg(jacksonimmunoresearch)。通過流式細(xì)胞術(shù)驗(yàn)證細(xì)胞耗盡的效率。
存活研究。死亡率用于評(píng)估急性lcmv誘導(dǎo)的腦膜炎的臨床嚴(yán)重性。小鼠每天檢查2次,共14天時(shí)間段,或直至達(dá)到100%的死亡率。
lcmv特異性足墊腫脹反應(yīng)的測定。小鼠如上所述在右后足墊中進(jìn)行局部感染,并且跟蹤局部腫脹反應(yīng)直至第14天p.i。用指針式測徑器(dialcaliper)(mitutoyo7309,mitutoyoco.,tokyo,日本)測量足墊厚度,并且病毒特異性腫脹測定為受感染右足和未感染左足厚度的差異(christensen等人,1994)。
細(xì)胞制備。無菌取出來自小鼠的脾細(xì)胞,并且轉(zhuǎn)移至hanks`平衡鹽溶液(hbss)。通過按壓器官經(jīng)由精細(xì)無菌鋼絲網(wǎng)獲得單細(xì)胞懸浮液。細(xì)胞用hbss洗滌2次,并且細(xì)胞濃度在包含10%胎牛血清(fcs)的rpmi1640中調(diào)整,所述rpmi1640補(bǔ)充有2-巰基乙醇、l-谷氨酰胺和青霉素-鏈霉素溶液。
用于流式細(xì)胞術(shù)的mab。下述mab都作為大鼠抗小鼠抗體購自pharmingen(sandiego,ca):fitc綴合的抗cd44、cy-鉻綴合的抗cd8a、cy-鉻綴合的抗cd4和藻紅蛋白(pe)綴合的抗ifn-γ。
流式細(xì)胞術(shù)分析。對(duì)于lcmv特異性(產(chǎn)生干擾素-γ)cd8+/cd4+t細(xì)胞的顯現(xiàn),使1-2x106個(gè)脾細(xì)胞重懸浮于0.2ml完全rpmi培養(yǎng)基中,所述完全rpmi培養(yǎng)基補(bǔ)充有10個(gè)單位鼠類重組il-2(r&dsystemseuropeltd,abingdon,uk)、3μm莫能星(sigmachemicalsco.,stlouis,mo)和1μg/ml相關(guān)肽,并且在37℃下孵育5小時(shí)。使用下述肽:對(duì)于cd8+t細(xì)胞為gp33-41、gp276-86、gp92-101、gp118-125且np396-404用于對(duì)照;對(duì)于cd4+t細(xì)胞為gp61-80。在孵育后,細(xì)胞進(jìn)行表面染色,洗滌,滲透化且用ifn-γ特異性mab染色,如先前描述的(andreasen等人,2000;christensen等人,2003)。同種型匹配的抗體充當(dāng)對(duì)照用于非特異性染色。細(xì)胞使用facscalibur(bectondickinson,sanjose,ca)進(jìn)行分析,并且使用低角度和側(cè)面散射的組合來門控至少104個(gè)活細(xì)胞,以排除死細(xì)胞和碎片。使用cell-quest軟件進(jìn)行數(shù)據(jù)分析。
實(shí)施例2:ii連接的抗原增強(qiáng)的cd8+t細(xì)胞激活不依賴于天然ii
ii序列包含在抗原加工具有功能的多個(gè)區(qū)域,其包括:細(xì)胞質(zhì)分選結(jié)構(gòu)域和三聚結(jié)構(gòu)域、細(xì)胞質(zhì)和近側(cè)膜發(fā)信號(hào)結(jié)構(gòu)域、細(xì)胞質(zhì)、膜內(nèi)和周質(zhì)三聚結(jié)構(gòu)域、涉及解鎖mhc分子以促進(jìn)外源肽結(jié)合的“key”基序、在clip區(qū)域中對(duì)于mhci和ii類的結(jié)合基序、周質(zhì)糖基化位點(diǎn)以及與cd44和巨噬細(xì)胞遷移抑制因子(mif)相互作用的在結(jié)構(gòu)上未鑒定的區(qū)域(圖2)。
ii連接增加在mhci和ii類上的抗原呈遞。通過使用骨髓衍生的樹突細(xì)胞(bmdc)的ad-iiova(ova是卵白蛋白)或ad-ova轉(zhuǎn)導(dǎo),我們發(fā)現(xiàn)ii連接的確誘導(dǎo)mhci類限制性抗原呈遞的顯著增加,如通過用針對(duì)在h-2kb上呈遞的siinfeklova表位的抗體的直接染色測量的(圖3)。增加的來自ii連接的序列的mhci類限制性抗原的表達(dá)直接作用于apc,不依賴于mhcii類、cd4+t細(xì)胞和任何其他細(xì)胞類型,并且可以在樹突細(xì)胞培養(yǎng)中直接測量。
以順式的ii
為了確定ii是僅以順式還是也以反式工作,通過合成具有β-珠蛋白多聚腺苷酸化信號(hào)的磷酸甘油酸酯激酶(pgk)啟動(dòng)子,并且將其克隆到腺病毒骨架的e3區(qū)域內(nèi),建立在腺病毒載體內(nèi)的另外讀碼框。這種載體隨后可以用于與穿梭載體重組,所述穿梭載體用于制備ad-gp載體(其表達(dá)在人cmv啟動(dòng)子和sv40多聚腺苷酸的控制下來自e1讀碼框的lcmvgp)。新載體表達(dá)來自腺病毒e1區(qū)的lcmvgp和來自e3區(qū)的ii(ad-ii+gp)。啟動(dòng)子通過轉(zhuǎn)染到cos7細(xì)胞內(nèi)就綠色熒光細(xì)胞的誘導(dǎo)進(jìn)行驗(yàn)證,并且在ad-iigp和ad-ii+gp感染的cos7細(xì)胞中iimrna表達(dá)的測量證實(shí)ii由pgk啟動(dòng)子表達(dá)至少與由cmv啟動(dòng)子表達(dá)一樣有效(圖4a)。用ad-gp、ad-iigp和ad-ii+gp感染的bmdc刺激的tcr318細(xì)胞的比較顯示ii必須與抗原連接,以具有任何作用(圖4b)。如果以反式的ii表達(dá)已顯示功效那將是令人驚訝的,因?yàn)橛糜诖碳さ腷mdc培養(yǎng)物已表達(dá)ii。
n末端改變:
從n末端開始,我們進(jìn)行1)前17個(gè)氨基酸的缺失(ad-δ17iigp),這去除基于亮氨酸的胞內(nèi)體分選信號(hào),2)用來自趨化因子受體ccr6tm6的相應(yīng)區(qū)段替換跨膜區(qū)段的前一半(ad-iiltmgp),3)用相應(yīng)ccr6tm6區(qū)段替換跨膜區(qū)段的另一半(ad-iiutmgp),和最后4)前50個(gè)氨基酸的完全缺失(ad-δ50iigp)。后面一種缺失去除整個(gè)細(xì)胞溶質(zhì)、tm和膜近側(cè)區(qū)。這些突變無一對(duì)其余ii序列增強(qiáng)cd8+t細(xì)胞刺激的能力具有任何作用(圖5)。
c末端改變:
從c末端我們進(jìn)行最后14個(gè)aa(ad-ii1-201gp,這去除c末端糖基化信號(hào))、最后97個(gè)aa(ad-ii1-118gp)和最后110個(gè)aa(ad-ii1-105gp)的缺失。通過1-201未觀察到對(duì)其余ii序列增強(qiáng)cd8+t細(xì)胞刺激的能力的作用,而從1-105突變和1-118突變僅可見不一致和較小的減少趨勢(圖6)。
突變:
我們還嘗試在報(bào)道的clip區(qū)域的mhci類結(jié)合位點(diǎn)(ad-iiclipgp:雙重的m至a點(diǎn)突變-m91am99a–設(shè)計(jì)為取消ii與mhci類分子的相互作用)和key基序(ad-iikeygp:lrmk至aaaa的四重的點(diǎn)突變,這將破壞ii-key區(qū)段)中進(jìn)行點(diǎn)突變。這些突變無一對(duì)于ii介導(dǎo)的增強(qiáng)的cd8+t細(xì)胞刺激是關(guān)鍵的。當(dāng)我們組合n和c末端截短(truncation)時(shí),唯一有趣的數(shù)據(jù)出現(xiàn)。因此,當(dāng)我們測試51-118變體(ad-ii51-118gp)時(shí),觀察到cd8+t細(xì)胞刺激能力的顯著減少,但突變體仍優(yōu)于ad-gp(圖7)。
實(shí)施例3
在本發(fā)明的一個(gè)實(shí)施方案中,提供了與ii偶聯(lián)的與糖基結(jié)合蛋白質(zhì)組合的非人糖基轉(zhuǎn)移酶。ii可以是全長或變體,其中變體可以是包括殘基編號(hào)50–215的ii的截短形式。這種變體具有完全活性,盡管缺乏跨膜結(jié)構(gòu)域。任選地,可以進(jìn)一步提供佐劑或一個(gè)或多個(gè)易位結(jié)構(gòu)域。在圖15中提供的是一個(gè)實(shí)施方案的示意圖,其中甘露糖受體(在疫苗中通常靶向的鈣依賴性凝集素)與包括殘基50–215的恒定鏈的變體(ii50-215)偶聯(lián),與腺病毒纖維蛋白質(zhì)進(jìn)一步偶聯(lián)。腺病毒纖維蛋白質(zhì)(ad纖維)可以源于腺病毒的任何血清型。甘露糖受體可以是來自甘露糖受體的一個(gè)或多個(gè)結(jié)構(gòu)域。
在一個(gè)特定例子中,提供了這樣的腺病毒:在一個(gè)讀碼框中表達(dá)egghead(來自果蠅的蛋白質(zhì)),并且在另一個(gè)讀碼框中表達(dá)甘露糖受體(或來自甘露糖受體的結(jié)構(gòu)域),所述受體與不含跨膜區(qū)的具有完全活性的ii變體例如ii50-215變體,其與腺病毒纖維蛋白質(zhì)進(jìn)一步偶聯(lián)。
糖基轉(zhuǎn)移酶例如egghead和糖基結(jié)合蛋白質(zhì)例如甘露糖受體可以由在相同腺病毒載體中的不同讀碼框表達(dá),或糖基轉(zhuǎn)移酶例如egghead和糖基結(jié)合蛋白質(zhì)例如甘露糖受體可以由同時(shí)施用的不同腺病毒載體表達(dá)。
egghead在所有糖基化er(內(nèi)質(zhì)網(wǎng))蛋白質(zhì)上偶聯(lián)甘露糖。分泌蛋白質(zhì)的甘露糖基化可以因此引起甘露糖化蛋白質(zhì)與甘露糖受體-ii-ad纖維復(fù)合物的結(jié)合(如圖15中所示)。復(fù)合物的腺病毒纖維引起與所述復(fù)合物連接的分泌蛋白質(zhì)由其他細(xì)胞吸收,激活這些以變得免疫刺激的,并且提供復(fù)合物對(duì)ii可以在其中發(fā)揮其作用的細(xì)胞溶質(zhì)的接近。
這種技術(shù)可以用于構(gòu)建可以直接施用到例如癌癥內(nèi)的疫苗。
參考文獻(xiàn)列表
andreasen,s.o.,christensen,j.e.,marker,o.&thomsen,a.r.(2000).roleofcd40ligandandcd28ininductionandmaintenanceofantiviralcd8+effectortcellresponses.jimmunol164,3689-3697.
bartholdy,c.,stryhn,a.,hansen,n.j.,buus,s.&thomsen,a.r.(2003).incompleteeffector/memorydifferentiationofantigen-primedcd8+tcellsingenegundna-vaccinatedmice.eurjimmunol33,1941-1948.
battegay,m.,cooper,s.,althage,a.,banziger,j.,hengartner,h.和zinkernagel,r.m.(1991).quantificationoflymphocyticchoriomeningitisviruswithanimmunologicalfocusassayin24-or96-wellplates.j.virol.methods33:191-198.
becker,t.c.,noel,r.j.,coats,w.s.,gomez-foix,a.m.,alam,t.,gerard,r.d.和newgard,c.b.(1994).useofrecombinantadenovirusformetabolicengineeringofmammaliancells.methodscellbiol.43pta:161-189.
christensen,j.p.,marker,o.&thomsen,a.r.(1994).theroleofcd4+tcellsincell-mediatedimmunitytolcmv:studiesinmhcclassiandclassiideficientmice.scandjimmunol40,373-382.
christensen,j.p.,kauffmann,s.o.&thomsen,a.r.(2003).deficientcd4+tcellprimingandregressionofcd8+tcellfunctionalityinvirus-infectedmicelackinganormalbcellcompartment.jimmunol171,4733-4741.
diebold,s.s.,m.cotten,n.koch和m.zenke.(2001).mhcclassiipresentationofendogenouslyexpressedantigensbytransfecteddendriticcells.genether.8:487-493.
holmesjp,benavideslc,gatesjd,carmichaelmg,huemanmt,mittendorfea,murrayjl,amina,craigd,vonhofee,ponniahs,peoplesge.(2008).resultsofthefirstphaseiclinicaltrialofthenovelii-keyhybridpreventiveher-2/neupeptide(ae37)vaccine.jclinoncol.jul10;26(20):3426-33.
holst,p.j.,sorensen,m.r.,mandrupjensen,c.m.,orskov,c.,thomsen,a.r.&christensen,j.p.(2008).mhcclassii-associatedinvariantchainlinkageofantigendramaticallyimprovescell-mediatedimmunityinducedbyadenovirusvaccines.jimmunol180,3339-3346.
kallinterisnl,lux,blackwellce,vonhofee,humphreysre,xum.(2006).ii-key/mhcclassiiepitopehybrids:astrategythatenhancesmhcclassiiepitopeloadingtocreatemorepotentpeptidevaccines.expertopin.biol.ther.6(12):1311-1321.
morris,c.r.,a.j.reber,j.l.petersen,s.e.vargas和j.c.solheim.(2004).associationofintracellularproteinswithfoldedmajorhistocompatibilitycomplexclassimolecules.immunol.res.30:171-179.
pieters,j.(1997).mhcclassiirestrictedantigenpresentation.curr.opin.immunol.9:89-96.
sambrook等人,編輯,molecularcloning:alaboratorymanual,coldspringharborlaboratory,2ndedition,coldspringharbor,n.y.,1989.
strumptner-cuvelette,p.和p.benaroch.(2002).multiplerolesoftheinvariantchaininmhcclassiifunction.biochem.biophys.acta.,1542:1-13.
wo2007/062656
us2008/0095798
wo2003/089471
us5,554,372
us5,876,735
5,102,985。
序列表
<110>holst,peterjohannes
thomsen,allanrandrup
christensen,janpravsgaard
grujic,mirjana
<120>免疫應(yīng)答的引發(fā)
<130>p1982dk00
<140>pa200801638
<141>2008-11-21
<160>4
<170>fastseqforwindowsversion4.0
<210>1
<211>1287
<212>dna
<213>智人-恒定鏈
<400>1
ctgcaggggggggggggggggggggggacattggctcttccttggggagtgatgcacagg60
aggagaagcaggagctgtcgggaagatcagaagccagtcatggatgaccagcgcgacctt120
atctccaacaatgagcaactgcccatgctgggccggcgccctggggccccggagagcaag180
tgcagccgcggagccctgtacacaggcttttccatcctggtgactctgctcctcgctggc240
caggccaccaccgcctacttcctgtaccagcagcagggccggctggacaaactgacagtc300
acctcccagaacctgcagctggagaacctgcgcatgaagcttcccaagcctcccaagcct360
gtgagcaagatgcgcatggccaccccgctgctgatgcaggcgctgcccatgggagccctg420
ccccaggggcccatgcagaatgccaccaagtatggcaacatgacagaggaccatgtgatg480
cacctgctccagaatgctgaccccctgaaggtgtacccgccactgaaggggagcttcccg540
gagaacctgagacaccttaagaacaccatggagaccatagactggaaggtctttgagagc600
tggatgcaccattggctcctgtttgaaatgagcaggcactccttggagcaaaagcccact660
gacgctccaccgaaagagtcactggaactggaggacccgtcttctgggctgggtgtgacc720
aagcaggatctgggcccagtccccatgtgagagcagcagaggcggtcttcaacatcctgc780
cagccccacacagctacagctttcttgctcccttcagcccccagcccctcccccatctcc840
caccctgtacctcatcccatgagaccctggtgcctggctctttcgtcacccttggacaag900
acaaaccaagtcggaacagcagataacaatgcagcaaggccctgctgcccaatctccatc960
tgtcaacaggggcgtgaggtcccaggaagtggccaaaagctagacagatccccgttcctg1020
acatcacagcagcctccaacacaaggctccaagacctaggctcatggacgagatgggaag1080
gcacagggagaagggataaccctacacccagaccccaggctggacatgctgactgtcctc1140
tcccctccagcctttggccttggcttttctagcctatttacctgcaggctgagccactct1200
cttccctttccccagcatcactccccaaggaagagccaatgttttccacccatccctccc1260
cccccccccccccccccccccctgcag1287
<210>2
<211>216
<212>prt
<213>智人-恒定鏈
<400>2
metaspaspglnargaspleuileserasnasngluglnleupromet
151015
leuglyargargproglyalaprogluserlyscysserargglyala
202530
leutyrthrglypheserileleuvalthrleuleuleualaglygln
354045
alathrthralatyrpheleutyrglnglnglnglyargleuasplys
505560
leuthrvalthrserglnasnleuglnleugluasnleuargmetlys
65707580
leuprolysproprolysprovalserlysmetargmetalathrpro
859095
leuleumetglnalaleuprometglyalaleuproglnglypromet
100105110
glnasnalathrlystyrglyasnmetthrgluasphisvalmethis
115120125
leuleuglnasnalaaspproleulysvaltyrproproleulysgly
130135140
serpheprogluasnleuarghisleulysasnthrmetgluthrile
145150155160
asptrplysvalpheglusertrpmethishistrpleuleupheglu
165170175
metserarghisserleugluglnlysprothraspalaproprolys
180185190
gluserleugluleugluaspproserserglyleuglyvalthrlys
195200205
glnaspleuglyprovalpromet
210215
<210>3
<211>648
<212>dna
<213>小家鼠-恒定鏈
<400>3
atggatgaccaacgcgacctcatctctaaccatgaacagttgcccatactgggcaaccgc60
cctagagagccagaaaggtgcagccgtggagctctgtacaccggtgtctctgtcctggtg120
gctctgctcttggctgggcaggccaccactgcttacttcctgtaccagcaacagggccgc180
ctagacaagctgaccatcacctcccagaacctgcaactggagagccttcgcatgaagctt240
ccgaaatctgccaaacctgtgagccagatgcggatggctactcccttgctgatgcgtcca300
atgtccatggataacatgctccttgggcctgtgaagaacgttaccaagtacggcaacatg360
acccaggaccatgtgatgcatctgctcacgaggtctggacccctggagtacccgcagctg420
aaggggaccttcccagagaatctgaagcatcttaagaactccatggatggcgtgaactgg480
aagatcttcgagagctggatgaagcagtggctcttgtttgagatgagcaagaactccctg540
gaggagaagaagcccaccgaggctccacctaaagagccactggacatggaagacctatct600
tctggcctgggagtgaccaggcaggaactgggtcaagtcaccctgtga648
<210>4
<211>215
<212>prt
<213>小家鼠-恒定鏈
<400>4
metaspaspglnargaspleuileserasnhisgluglnleuproile
151015
leuglyasnargproarggluprogluargcysserargglyalaleu
202530
tyrthrglyvalservalleuvalalaleuleuleualaglyglnala
354045
thrthralatyrpheleutyrglnglnglnglyargleuasplysleu
505560
thrilethrserglnasnleuglnleugluserleuargmetlysleu
65707580
prolysseralalysprovalserglnmetargmetalathrproleu
859095
leumetargprometsermetaspasnmetleuleuglyprovallys
100105110
asnvalthrlystyrglyasnmetthrglnasphisvalmethisleu
115120125
leuthrargserglyproleuglutyrproglnleulysglythrphe
130135140
progluasnleulyshisleulysasnsermetaspglyvalasntrp
145150155160
lysilepheglusertrpmetlysglntrpleuleupheglumetser
165170175
lysasnserleugluglulyslysprothrglualaproprolysglu
180185190
proleuaspmetgluaspleuserserglyleuglyvalthrarggln
195200205
gluleuglyglnvalthrleu
210215
- 還沒有人留言評(píng)論。精彩留言會(huì)獲得點(diǎn)贊!
- 細(xì)胞介導(dǎo)的免疫應(yīng)答試驗(yàn)的制作方法
- 鳥類中增強(qiáng)的免疫應(yīng)答的制作方法
- 免疫應(yīng)答的增強(qiáng)的制作方法
- 用于選擇和去選擇用產(chǎn)生免疫應(yīng)答的療法治療的癌癥患者的質(zhì)譜方法
- 應(yīng)答裝置的制造方法
- 調(diào)節(jié)免疫應(yīng)答的制作方法
- 折疊式便攜信息終端、用于其中的應(yīng)答保持方法及其程序的制作方法
- 電話、來訪兩用應(yīng)答錄音裝置的制作方法
- 模子生產(chǎn)過程中所用基片的一種制造方法及利用該方法得到的一種基片的制作方法
- 使用連接寡核苷酸調(diào)節(jié)tlr介導(dǎo)的免疫應(yīng)答的制作方法
- 用于減弱外顯子跳讀抗病毒轉(zhuǎn)移載體免疫應(yīng)答的方法和組合物與流程
- 用于減弱基因治療抗病毒轉(zhuǎn)移載體免疫應(yīng)答的方法和組合物與流程
- 免疫應(yīng)答的引發(fā)的制造方法與工藝
- 用于增強(qiáng)免疫應(yīng)答的方法和組合物與流程
- 在有需要的對(duì)象中刺激免疫應(yīng)答的方法與流程
- 表達(dá)免疫應(yīng)答刺激細(xì)胞因子以吸引和/或激活免疫細(xì)胞的基因修飾間充質(zhì)干細(xì)胞的制造方法與工藝
- 一種研究北蟲草多糖對(duì)機(jī)體的體液免疫應(yīng)答和細(xì)胞免疫應(yīng)答反應(yīng)的方法與制造工藝
- 用于減弱基因治療抗病毒轉(zhuǎn)移載體免疫應(yīng)答的方法和組合物與流程
- 免疫應(yīng)答的引發(fā)的制造方法與工藝
- 用于增強(qiáng)免疫應(yīng)答的方法和組合物與流程
- 在有需要的對(duì)象中刺激免疫應(yīng)答的方法與流程
- 表達(dá)免疫應(yīng)答刺激細(xì)胞因子以吸引和/或激活免疫細(xì)胞的基因修飾間充質(zhì)干細(xì)胞的制造方法與工藝
- 一種研究北蟲草多糖對(duì)機(jī)體的體液免疫應(yīng)答和細(xì)胞免疫應(yīng)答反應(yīng)的方法與制造工藝
- 用于增強(qiáng)免疫應(yīng)答的泰帕森(Tapasin)增加的制作方法
- 免疫應(yīng)答的引發(fā)的制作方法