一種夾氧雜蒽酮類化合物CCE9的用途的制作方法
本發明屬于中藥提取物的應用領域,具體涉及一種夾氧雜蒽酮類化合物cce9的用途。
背景技術:
p38絲裂原活化蛋白激酶(p38mitogen-activatedproteinkinases(p38mapk))可以由細胞外的多種應急包括uv,放射線,熱休克,促炎癥分子,特定抗原及其它應激反應激活,并在凋亡,細胞因子產生,轉錄調節及癲癇發作中其重要的作用。
近期研究表明p38mapk在細胞凋亡中起重要作用。如p38mapk通路的激活可導致神經細胞凋亡。在腫瘤細胞中,p38mapk活性升高,并參與調控凋亡。最近的研究表明p38mapk可以通過誘導bcl-2磷酸化,誘導細胞凋亡。
bcl-2是細胞凋亡過程最重要的調控因子之一。最近,越來越多的證據表明,bcl-2的結構無序鏈區域的磷酸化調控著bcl-2的生存功能,bcl-2磷酸化失活其生存功能。在環境因子、炎癥細胞因子、脂多糖(lps)和其它因子的作用下,p38mapk可以介導bcl-2結構無序鏈上ser87和thr56的磷酸化,從而介導ngf撤除、腫瘤壞死因子和no所誘導的細胞凋亡過程。在上述情況下,無論是細胞中或無細胞實驗中,p38mapk介導的bcl-2高度磷酸化均表現為促凋亡事件,誘導細胞色素c從線粒體的釋放過程。
bcl-2結構無序鏈區域的磷酸化可能通過影響其與bcl-2家族促凋亡蛋白如bax的相互作用抑制其抗凋亡功能。在記憶性b淋巴細胞中,ngf撤除誘導依賴p38mapk的磷酸化,抑制其抗凋亡功能,誘導細胞色素c的釋放。同樣,在成年大鼠心臟細胞中h2o2介導依賴p38mapk的bcl-2磷酸化,誘導細胞凋亡。已有證據表明,bcl-2結構無序鏈上的ser87和thr56是p38mapk磷酸化bcl-2的位點,并且這此殘基的磷酸化降低bcl-2的抗凋亡作用。
cce9是一種從中藥牛黃木中提取的夾氧雜蒽酮類化合物。黃牛木(cratoxyloncochinchinensisbl)別稱黃牛茶、黃芽木、狗芽木、雀籠木、水杧果、鷓鴣木、節節花、滿天紅(廣西)、茶咯桌、梅低優、美啟烈、山狗芽等是藤黃科(guttiferae)黃牛木屬(cratoxylum)植物。功用清熱解暑、化濕消滯、消腫止血,主治感冒發熱、腸炎腹瀉、咳嗽聲嘶、黃疸病等;黃牛木的主要化學成分為夾氧雜蒽酮類,此外還含有蒽醌苯甲酮類、三萜類和黃酮類成分;現代藥理研究表明黃牛木具有抗氧化、細胞毒性和神經生長因子增效作用。
雖然已有文獻報道黃牛木具有抗癌活性,并分離得到了一些活性成分,但研究大多只限于較為基礎的mtt實驗階段如laphookhieo等對c.cochinchinense根中分離到的7個吡酮類化合物進行體外細胞毒活性測試,結果與喜樹堿的抗癌活性比較表明化合物cochinchinonea對乳腺癌,宮頸癌,腸癌和kb細胞系具有較強的生長抑制活性;boonnaknawong等發現化合物gerontoxanthonei對乳腺癌,宮頸癌,腸癌和kb細胞系具有較強的細胞毒活性等,但是這些簡單的活性研究無法明確活性成分的抗腫瘤作用靶點、作用機理,這是制約其進一步開發成為抗癌藥物的最主要原因。
技術實現要素:
本發明的第一個目的在于,提供一種夾氧雜蒽酮類化合物cce9在制備治療癌癥藥物中的用途。
本發明的第二個目的在于,提供一種以bcl-2作為作用靶點篩選夾氧雜蒽酮類化合物的應用。
為了實現本發明目的,本發明提供了一種夾氧雜蒽酮類化合物cce9用于制備治療癌癥藥物的用途。
進一步,所述夾氧雜蒽酮類化合物cce9以bcl-2為作用靶點。
進一步,所述夾氧雜蒽酮類化合物cce9具有用于誘導腫瘤細胞凋亡的用途。
進一步,所述夾氧雜蒽酮類化合物cce9具有通過激活p38mapk誘導腫瘤細胞凋亡的用途。
進一步,所述夾氧雜蒽酮類化合物cce9具有通過誘導bcl-2的磷酸化誘導腫瘤細胞凋亡的用途。
進一步,所述夾氧雜蒽酮類化合物cce9具有通過誘導bcl-2的構象變化誘導腫瘤細胞凋亡的用途。
進一步,所述夾氧雜蒽酮類化合物cce9具有通過bax的激活誘導腫瘤細胞凋亡的用途。
進一步,所述癌癥為子宮頸癌、肺癌和肝癌。
所述夾氧雜蒽酮類化合物cce9(1,3,7-trihydroxy-2,4-diprenylxanthone)為中藥黃牛木提取物,其結構如下:
根據本發明,以bcl-2的磷酸化為靶點,可以用于篩選夾氧雜蒽酮類化合物,以進一步開發用于治療各種癌癥。
本發明提供的作用于bcl-2誘導腫瘤細胞凋亡可以用于指導篩選夾氧雜蒽酮類化合物,從中獲得具有抗腫瘤活性的化合物,并可將獲得的藥物進而制備可用于bcl-2誘導腫瘤細胞凋亡的活性與功能的藥物。
附圖說明
圖1a為熒光染色檢測cce9誘導腫瘤細胞的凋亡圖;
圖1b為cce9誘導腫瘤細胞凋亡的百分比結果圖;
圖1c為cce9誘導腫瘤細胞中的parp蛋白切割圖;
圖2為流式細胞儀分析annexin-v/pi雙染檢測cce9誘導hela229子宮頸癌細胞的凋亡結果圖;
圖3為cce9對hela229細胞中p38mapk的激活作用圖;
圖4a為p38mapk抑制劑抑制cce9誘導的腫瘤細胞凋亡圖;
圖4b為敲低p38表達抑制cce9誘導的腫瘤細胞凋亡圖;
圖5為流式細胞儀分析annexin-v/pi雙染檢測敲除細胞中的bcl-2抑制cce9誘導的腫瘤細胞凋亡圖;
圖6為cce9對hela229細胞中bcl-2的磷酸化作用以及p38mapk抑制劑對bcl-2的磷酸化的抑制作用圖;
圖7為cce9誘導hela229細胞中bcl-2的構象變化圖;
圖8為cce9誘導hela229細胞中bax的激活圖;
圖9a為p38mapk抑制劑抑制bcl-2的構象變化圖;
圖9b為敲低p38表達抑制cce9誘導的bcl-2的構象變化圖。
具體實施方式
下面詳細描述本發明的實施例,所述實施例的示例旨在用于解釋本發明,而不能理解為對本發明的限制。實施例中未注明具體條件的實驗方法,通常按照常規條件如《分子克隆:實驗室手冊》(newyork:coldspringharborlaboratorypress,1989)中所述的條件,或按照制造廠商所建議的條件。
本發明中使用的試劑:
所采用的夾氧雜蒽酮類化合物cce9其結構如下:
實施例1-4、6-9中所述10μmcce9溶液由cce9溶解于99.9%dmso溶液(dmso終濃度<0.1%),配制10mmcce9溶液,用1μlcce9溶液加入1mlmem培養基中配制得到。
實施例5中所述10μmcce9溶液由cce9溶解于99.9%dmso溶液(dmso終濃度<0.1%),配制10mmcce9溶液,用1μlcce9溶液加入1mldmem培養基中配制得到。
實施例1cce9誘導腫瘤細胞凋亡
1、細胞培養
選擇子宮頸癌細胞株hela229(atcc),采用10%小牛血清的mem培養基,在恒溫37℃、5%co2培養箱中取出培養于24孔組織培養板中,24小時后換液和進行加藥(不含血清)處理。cce9溶解于99.9%dmso溶液(dmso終濃度<0.1%),配制10mmcce9溶液,用1μlcce9溶液加入1mlmem培養基中,cce9終濃度為10μm,處理細胞3小時和6小時,對照組用同樣濃度dmso溶液處理6小時。
2、dapi染色細胞核
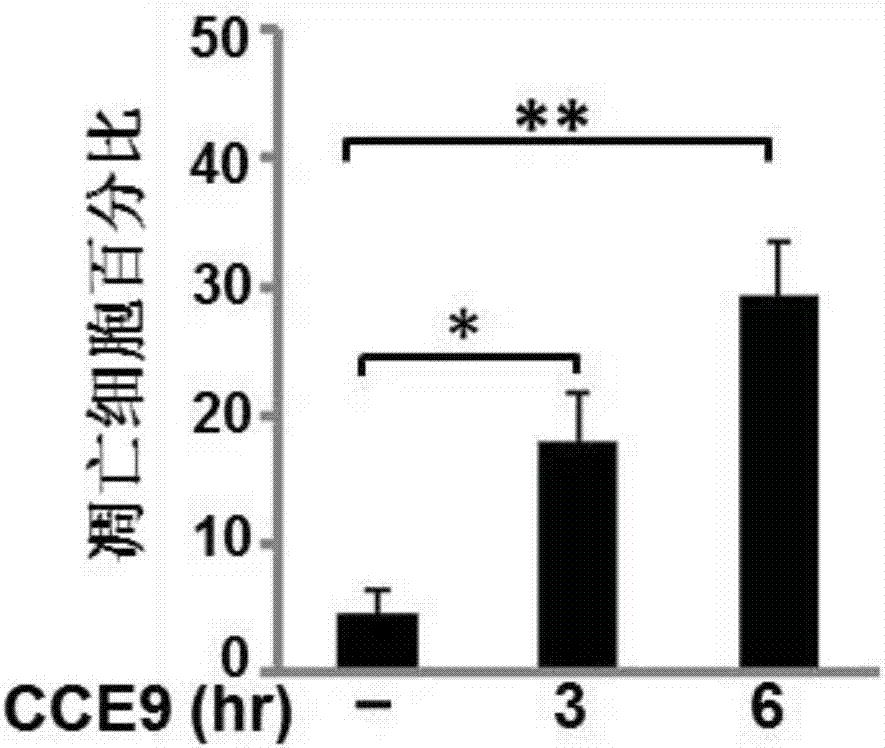
反應板中各孔溶液的總體積均為1ml。細胞經處理后,pbs洗3次,4%多聚甲醛固定,1%triton穿膜,最后加入50μg/mldapi加入100μg/mldnase-freernasea37℃染細胞核20分鐘。甘油封片,熒光顯微鏡下觀察細胞核形態,結果如圖1a所示。至少300個細胞,多于5個隨機視然野,由兩個不同的觀察者記數,計算凋亡的百分比,結果如圖1b所示。
由圖1a可見,用10μm的cce9溶液作用hela229細胞3小時和6小時,作用3小時的hela229細胞明顯發生凋亡,作用6小時的hela229細胞數量比作用3小時的hela229細胞更少,且細胞質更皺,凋亡細胞根據形態上典型的細胞質皺縮,膜起泡以及核凝縮、破碎加以識別。
由圖1b統計結果顯示,在對照組hela229細胞極少發生凋亡的情況下(4.7%),作用cce9溶液3小時后凋亡率達到17%,且隨著作用時間的延長,6小時后凋亡hela229細胞達到34.3%,hela229細胞凋亡顯著增加。
3、cce9誘導hela229細胞、a549細胞和hepg2細胞的papr切割
聚腺苷酸二磷酸核糖轉移酶(poly(adp-ribose)polymerase,parp)的切割是早期凋亡的標記。parp的切割可以通過其抗體加以識別。選擇子宮頸癌細胞株hela229(atcc),用10%小牛血清的mem培養基,肺癌細胞株a549(atcc)和肝癌細胞株hepg2(atcc),用10%小牛血清的dmem培養基,恒溫37℃、5%co2培養箱中取出培養于12孔組織培養板中,24小時后換液和進行加藥(不含血清)處理。cce9溶解于99.9%dmso溶液(dmso終濃度<0.1%),配制10mmcce9溶液,用1μlcce9溶液加入1mlmem培養基中,cce9終濃度為10μm,處理0.25、0.5、1、3小時和6小時,對照組用同樣濃度dmso處理6小時。細胞以ripa細胞裂解液(50mmtris-hclph7.4,1%np-40,0.25%na-deoxycholate,150mmnacl)裂解30分鐘,以相同上樣量,8%sds-page電泳,轉膜,以5%脫脂奶粉的tbst(50mmtris-hcl(ph7.4),150mmnacland0.1%tween-20)室溫封閉1小時,4℃過夜孵育一抗anti-parp(1:1000稀釋),室溫孵育二抗1小時,ecl顯色,曝光。結果如圖1c所示。
如圖1c所示,10μmcce9溶液作用3小時,就可引起子宮頸癌細胞株hela229中parp的切割;a549肺癌細胞在10μmcce9溶液作用6小時,發生parp切割;hepg2肝癌細胞在10μmcce9溶液作用1小時中可觀察到了同樣的parp切割作用。
由圖1a、圖1b、圖1c可得,cce9可誘發子宮頸癌細胞、肝癌細胞和肺癌細胞明顯發生凋亡。
實施例2annexin-v/pi雙染檢測cce9誘導hela229子宮頸癌細胞的凋亡
在無血清mem(購買自hyclone)培養基中,用10μmcce9溶液作用hela229細胞6小時,以不加cce9溶液的細胞為對照。細胞根據vybrantapoptosisassaykit#2操作手冊染色pi和annexin-v,用流式細胞儀(beckmancoultercytoflex)分析。用beckmancoultercytexpert軟件分析,結果如圖2所示。
由圖2可以看出,對照組中只有1.87%的hela229細胞早期凋亡,而用10μmcce9溶液處理的實驗組有19.85%的hela229細胞早期凋亡。因此可得,cce9可誘發hela229細胞明顯發生凋亡。
實施例3cce9對hela229細胞中p38mapk的激活作用
1、細胞培養
選擇子宮頸癌細胞株hela229(atcc),用10%小牛血清的mem培養基,恒溫37℃、5%co2培養箱中取出培養于12孔組織培養板中,24小時后換液和進行加藥(不含血清)處理。cce9溶解于99.9%dmso溶液(dmso終濃度<0.1%),配制10mmcce9溶液,用1μlcce9溶液加入1mlmem培養基中,使得cce9終濃度為10μm,以10μmcce9溶液處理0.25、0.5、1、3小時和6小時,對照組用同樣濃度dmso處理6小時。
2、westernblotting分析p38mapk的激活
細胞以ripa細胞裂解液(50mmtris-hclph7.4,1%np-40,0.25%na-deoxycholate,150mmnacl)裂解30分鐘,以相同上樣量,10%sds-page電泳,轉膜,以5%脫脂奶粉的tbst(50mmtris-hcl(ph7.4),150mmnacland0.1%tween-20)室溫封閉1小時,4℃過夜孵育一抗anti-p-p38(1:1000稀釋),室溫孵育二抗1小時,ecl顯色,曝光。結果如圖3所示。
如圖3所示,從左往右的泳道依次代表cce9溶液處理hela229細胞的時間分別為0h,0.25h,0.5h,1h,3h,6h。由0.25h對應的泳道開始出現條帶,且隨著時間的增加條帶顏色越來越深,說明10μmcce9溶液作用25分鐘,就可引起子宮頸癌細胞株hela229中p38的激活,且隨作用時間的增加,激活程度逐漸增強。因此,cce9能夠快速而且有效的激活p38mapk,且這種激活都呈時間依賴型的規律。
實施例4cce9對hela229細胞的凋亡作用依賴于p38mapk的激活
1、p38mapk抑制劑sb203580抑制cce9誘導的腫瘤細胞凋亡
在無血清mem(購買自hyclone)培養基中,在p38mapk抑制劑sb203580、sp600125存在、缺少的情況下,用10μmcce9溶液作用hela229細胞3小時,以不加cce9溶液的細胞為對照。westernblotting分析parp的切割,方法同實施例1中3的實驗方法,結果如圖4a所示。
如圖4a所示:不進行任何處理的對照組、只加sb203580實驗組、同時加入sb203580和cce9溶液處理的實驗組p-p38對應的泳道未出現條帶,只加入sp600125的實驗組中p-p38對應的泳道出現一小段條帶,只加入cce9溶液和同時加入cce9溶液與sp600125的實驗組中p-p38對應的泳道出現長且粗的條帶。由此可得。只加入cce9溶液和同時加入cce9溶液與sp600125的實驗組中很多parp切割;而只加入sb203580和同時加入cce9溶液與sb203580的實驗組parp切割的量很少;只加入sp600125的實驗組中parp切割的量比只加入sb203580多。由圖4a可得,sb203580可以很大程度的抑制cce9誘導的parp切割,而sp600125(jnk抑制劑)只具有微弱的影響。
2、敲低p38表達抑制cce9誘導的腫瘤細胞凋亡
在hela229細胞中轉入p38mapk的特異sirna(5’-cugagaaacauauugugaudtdt-3’;5’-cuuguaagaucacucuuaadtdt-3’;5’-gaagcaaugggaauuuacadtdt-3’,購自sigma)48小時,用10μmcce9溶液作用hela229細胞3小時,以不加cce9溶液的細胞為對照。westernblotting分析parp的切割,方法同實施例1中3的實驗方法,結果如圖4b所示。
如圖4b所示,在hela229細胞中轉入p38mapk的特異sirna的實驗組中p38mapk的不表達,無parp切割。因此可得降低細胞中p38mapk的表達能夠顯著的抑制cce9誘導的腫瘤細胞中的parp切割。
以上結果均說明p38mapk在cce9誘導的腫瘤細胞凋亡中發揮重要的作用。
實施例5流式細胞儀分析annexin-v/pi雙染檢測敲除細胞中的bcl-2抑制cce9誘導的腫瘤細胞凋亡
在無血清dmem(購買自hyclone)培養基中,用10μmcce9溶液分別作用mef細胞和mefbcl-2敲除細胞(bcl-2-/-mef)6小時,以不加cce9溶液的細胞為對照。細胞根據vybrantapoptosisassaykit#2操作手冊染色pi和annexin-v,用流式細胞儀(beckmancoultercytoflex)分析。用beckmancoultercytexpert軟件分析,結果如圖5所示,mef細胞的對照組2.8%細胞早期凋亡,mef細胞加cce9溶液處理的實驗組16.51%的細胞早期凋亡;mefbcl-2敲除細胞的對照組3.62%細胞早期凋亡,mefbcl-2敲除細胞加cce9溶液處理的實驗組3.73%的細胞早期凋亡。
因此由圖5可得,在mefbcl-2敲除細胞(bcl-2-/-mef)中,cce9并不能誘導細胞發生凋亡。
實施例6cce9對hela229細胞中bcl-2的磷酸化作用以及p38mapk抑制劑對bcl-2的磷酸化的抑制作用
在無血清mem(購買自hyclone)培養基中,在p38mapk抑制劑sb203580存在、缺少的情況下,用10μmcce9溶液作用hela229細胞3小時,以不加cce9溶液的細胞為對照。細胞以ripa細胞裂解液(50mmtris-hclph7.4,1%np-40,0.25%na-deoxycholate,150mmnacl)裂解30分鐘,加入bcl-2的抗體(購買自santacruz)10μl,于4℃搖床孵育2小時,然后加入30μlproteina/gagarose,4℃搖床孵育1-2小時;然后4℃2000轉離心2分鐘,小心的洗去上清,再用冰涼的細胞裂解液1ml在4℃搖床上洗滌至少3次,每次5分鐘,最后在4℃2000轉離心2分鐘,收集珠子。向離心管中加入適量的2×sdsloadingbuffer于100℃中變性10分鐘。以相同上樣量,12%sds-page電泳,轉膜,以5%脫脂奶粉的tbst(50mmtris-hcl(ph7.4),150mmnacland0.1%tween-20)室溫封閉1小時,4℃過夜孵育一抗anti-p-thr(1:1000稀釋)或anti-p-ser(1:1000稀釋),室溫孵育二抗1小時,ecl顯色,曝光。結果如圖6所示。
如圖6所示,cce9可以誘導bcl-2依賴于絲氨酸和蘇氨酸的磷酸化,而p38mapk的抑制劑sb203580可以抑制cce9誘導的bcl-2磷酸化。從而,cce9能夠誘導依賴于p38mapk的bcl-2磷酸化。
實施例7cce9誘導hela229細胞中bcl-2的構象變化
在無血清mem培養基中,用10μmcce9溶液作用hela229細胞2小時,以不加cce9溶液的細胞為對照。細胞用4%甲醛(pbs中)室溫固定10分鐘,0.1%tritonx-100+0.1mgycine冰上穿膜30分鐘,5mg/mlbsa室溫封閉1小時,一抗anti-bcl-2(bh3),(用5mg/mlbsa稀釋1:50-100)(購自abcom)一個特異識別bcl-2的bh3結構域的抗體,只有bcl-2的構像發生改變后才能夠被bcl-2(bh3)抗體所識別,在濕盒中室溫孵育1-3小時,二抗(5mg/mlbsa稀釋1:50-100)濕盒中室溫黑暗中孵育1-3小時,dapi(1μg/mlinpbs)室溫孵育1-5分鐘,封片,在lsm-510共聚焦顯微鏡(carlzeiss,oberkochen,germany)觀察,并拍照。結果如圖7所示。
如圖7所示,雖然hela229細胞中有高表達的bcl-2蛋白,但對照組細胞中的bcl-2無法被bcl-2(bh3)抗體染色,說明hela229胞中bcl-2的bh3結構域隱藏在蛋白分子的內部。細胞經cce9處理后,呈顯bcl-2(bh3)抗體染色陽性。說明cce9可以誘導bcl-2構象改變。
實施例8cce9誘導hela229細胞中bax的激活
在無血清mem培養基中,用10μmcce9溶液作用hela229細胞2小時,以不加cce9溶液的hela229細胞為對照。如實施例7所述方法,用anti-bax(6a7),一個特異識別激活bax的抗體,染色。在lsm-510共聚焦顯微鏡(carlzeiss,oberkochen,germany)觀察,并拍照。結果如圖8所示。
如圖8所示,雖然hela229細胞中有高表達的bax蛋白,但對照組hela229細胞中的bax無法被bax(6a7)抗體染色,cce9處理引起hela229細胞中bax(6a7)的強染色,說明cce9可以激活bax。
實施例9cce9誘導的bcl-2的構象變化依賴于p38mapk的激活
1、p38mapk抑制劑抑制bcl-2的構象變化
在無血清mem(購買自hyclone)培養基中,在p38mapk抑制劑sb203580存在、缺少的情況下,用10μmcce9溶液作用hela229細胞2小時,以不加cce9溶液的細胞為對照。實驗方法同實施例7,免疫熒光染色法檢測bcl-2的構象改變情況,結果如圖9a所示。
如圖9a所示:雖然hela229細胞中有高表達的bcl-2蛋白,但對照組hela229細胞中的bcl-2無法被bcl-2(bh3)抗體染色,hela229細胞經cce9處理后,呈顯bcl-2(bh3)抗體染色陽性,而同時經cce9和sb203580處理的hela229細胞呈顯bcl-2(bh3)抗體染色陰性,由此可得sb203580可以很大程度的抑制cce9誘導的bcl-2的構象變化。
2、敲低p38表達抑制cce9誘導的bcl-2的構象變化
在hela229細胞中轉入p38mapk的特異sirna48小時,用10μmcce9溶液作用hela229細胞3小時,以不加cce9溶液的細胞為對照。用實施例7所述方法,免疫熒光染色法檢測bcl-2的構象改變情況,結果如圖9b所示。
如圖9b所示:雖然hela229細胞中有高表達的bcl-2蛋白,但對照組hela229細胞中的bcl-2無法被bcl-2(bh3)抗體染色,hela229細胞經cce9處理后,呈顯bcl-2(bh3)抗體染色陽性,而同時經cce9處理和轉入p38mapk的特異sirna的細胞呈顯bcl-2(bh3)抗體染色陰性。由此可得,降低hela229細胞中p38mapk的表達能夠顯著的抑制cce9誘導的bcl-2的構象變化。
以上結果均說明p38mapk在cce9誘導的bcl-2的構象變化中發揮重要的作用。
綜上所述,本發明的夾氧雜蒽酮類化合物cce9通過激活p38mapk,使得bcl-2發生磷酸化,導致bcl-2構象改變,激活bax,從而誘導腫瘤細胞的凋亡。因此,夾氧雜蒽酮類化合物cce9可以用于制備治療癌癥的以bcl-2為作用靶點的藥物。另外,bcl-2磷酸化可以作為作用靶點用于篩選夾氧雜蒽酮類化合物。
盡管上面已經示出和描述了本發明的實施例,可以理解的是,上述實施例是示例性的,不能理解為對本發明的限制,本領域的普通技術人員在不脫離本發明的原理和宗旨的情況下在本發明的范圍內可以對上述實施例進行變化、修改、替換和變型。
- 還沒有人留言評論。精彩留言會獲得點贊!