抗人EGFR和Notch多特異性抗體、其制備方法及用途與流程
本發明屬于生物醫藥技術領域,更具體地,本發明公開了一類抗人EGFR、Notch2和Notch3雙特異性抗體、其制備方法及其在制備抗腫瘤藥物中的應用。
背景技術:
以手術治療、放射治療及化學治療為主,輔以新型的靶向治療方案是近年來對惡性腫瘤治療的基本策略,且已經在臨床實踐中取得重要的進展。但惡性腫瘤的復發、轉移和治療性耐受依然是一直困擾臨床和科研工作者的難題。2006年,在美國癌癥研究協會召開的腫瘤干細胞研討會上,與會學者定義腫瘤干細胞為一種具有自我更新特性、有能力驅動形成多種異質性的腫瘤分化細胞群、具有干細胞特性的癌細胞(Clarke et al.,Cancer stem cells—perspectives on current status and future directions:AACR Workshop on cancer stem cells.,2006,Cancer research,66:9339-9344)。腫瘤干細胞學說以其獨特的理論優勢和不斷豐富的研究支持證據,越來越受到國內外學者的認同和接受。傳統治療方法雖然對腫瘤實體細胞有較好的治療殺傷效果,但對只占腫瘤細胞群0.2%~10%、并與腫瘤實體細胞具有較大分子表型差異的腫瘤干細胞難有抑制作用。研究腫瘤干細胞在腫瘤產生、發展和轉移的重要作用及其機理,以及開發新型的基于腫瘤干細胞學說的抗腫瘤治療策略及制劑,對惡性腫瘤的治療具有十分重要的意義。
腫瘤干細胞學說認為:惡性腫瘤起源于一群“干細胞樣”的腫瘤細胞,這群細胞被稱為腫瘤干細胞(cancer stem cells,CSCs or cancer-initiating cells,CICs)。上世紀90年代末,研究人員首先從白血病細胞中分離到腫瘤干細胞并闡述了由腫瘤干細胞驅動的腫瘤異質性分層組織模型(Dick,Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell.,1997,Nature Med,3:730-737)。隨后,研究人員在多種實體瘤中也證實了腫瘤干細胞的存在和類似的腫瘤異質性組織特點,包括乳腺癌,結腸癌,腦腫瘤,卵巢癌,肺癌,前列腺癌及胰腺癌(Brooks et al.,Therapeutic implications of cellular heterogeneity and plasticity in breast cancer.,2015,Cell stem cell,17:260-271)。這些證據均表明在大部分血液腫瘤和實體瘤中存在具有干細胞特性的腫瘤細胞。
多項研究顯示,腫瘤干細胞在惡性腫瘤發生、發展;腫瘤細胞異質性維持;腫瘤進化;和腫瘤治療性耐受的過程中發揮重要的作用。其生物學功能具有以下特點:
1)極強的致瘤性。從惡性腫瘤患者的癌灶中直接分離的腫瘤細胞很難在體外形成克隆或移植至免疫缺陷老鼠模型中進行克隆成瘤,而只需要少量的CSC就可以在免疫缺陷小鼠模型中克隆成瘤,且腫瘤移植物具有和腫瘤原發灶類似的表型異質性(Al-Hajj et al.,Prospective identification of tumorigenic breast cancer cells.,2003,Proceedings of the National Academy of Sciences,100:3983-3988;Dick,Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell.,1997,Nature Med,3:730-737;Ginestier et al.,ALDH1is a marker of normal and malignant human mammary stem cells and a predictor of poor clinical outcome.,2007,Cell stem cell,1:555-567)。
2)復雜的異質性及生物可塑性。CSC的來源可能及其復雜,其可能來源于在上皮細胞向間充質細胞轉化(epithelial-to-mesenchymal transition,EMT)的過程中獲得了自我更新能力的已分化的腫瘤細胞(Espinoza and Miele,Deadly crosstalk:Notch signaling at the intersection of EMT and cancer stem cells.,2013,Cancer letters,341:41-45);或者來源于由外來微環境因子誘導癌基因性突變的正常組織干細胞(Reya et al.,Stem cells,cancer,and cancer stem cells.,2001,nature,414:105-111)。另外,近年來的一些報道也證實CSC具有非常復雜的生物可塑性,如在乳腺癌干細胞中可以主要鑒定出兩種不同表型的乳腺癌干細胞群:一種上皮樣的增生活躍的乳腺癌CSC群,表達干細胞標志物ALDH;和一種間質樣的、相對不活躍但浸潤能力強的乳腺癌CSC群,其表面標志物表達情況為CD44+/CD24-。這兩群CSC都能產生各自的上皮樣或間質樣腫瘤實體細胞子代,同時可以釋放分泌信號增強CSC的自我更新能力。且在受到腫瘤微環境中細胞因子信號、趨化因子信號、轉錄調控介導的表觀遺傳學改變下,兩群細胞可以相互轉化(Liu et al.,Breast cancer stem cells transition between epithelial and mesenchymal states reflective of their normal counterparts.,2014,Stem cell reports,2:78-91)。另外,研究也報道在肝癌中存在不同CSC群,如CD133+的CSC(Ma et al.,CD133+HCC cancer stem cells confer chemoresistance by preferential expression of the Akt/PKB survival pathway.2008,Oncogene,27:1749-1758)和CD90+/CD44+的肝癌CSC(Yang et al.,Significance of CD90+cancer stem cells in human liver cancer.,2008,Cancer cell,13:153-166)具有不同的干細胞特性。不同的腫瘤類型中干細胞的標志物也往往具有差異,如在胰腺癌中,CSC的表面標記物被認為是CD24+(Li et al.,Structural basis for inhibition of the epidermal growth factor receptor by cetuximab.,2007,Cancer research,67:1030-1037),而對于乳腺CSC,則為CD24-。另外,不同惡性腫瘤中CSC的含量及比率往往不同,且在惡性腫瘤的不同階段,CSC的比率也會發生復雜的變化。如在結腸癌中,CD133+高表達CSC比率約為1.8%~24.5%(O’Brien et al.,A human colon cancer cell capable of initiating tumour growth in immunodeficient mice.,2007, Nature,445:106-110),而在接受化療后,結腸癌CSC的比率則會提高(Dylla et al.,Colorectal cancer stem cells are enriched in xenogeneic tumors following chemotherapy.,2008,PloS one,3:e2428)。而在黑色素瘤中,表達CD133+細胞表面標記的CSC的比率在1%~20%之間(Quintana et al.,Efficient tumour formation by single human melanoma cells.,2008,Nature,456:593-598)。這些研究結果均提示CSC具有極其復雜的異質性及生物可塑性。
3)治療敏感性差。近年來的一些研究表明,CSC對許多目前正在臨床中使用的治療方法耐受,包括多種化療藥物和放射療法。如有報道顯示人急性髓系白血病CD34+/CD38-祖細胞對化療殺傷作用及Fas介導的凋亡作用不敏感(Costello et al.,Human acute myeloid leukemia CD34+/CD38- progenitor cells have decreased sensitivity to chemotherapy and Fas-induced apoptosis,reduced immunogenicity,and impaired dendritic cell transformation capacities.,2000,Cancer Research,60:4403-4411);且高表達端粒酶,維持染色體的正常結構從而抑制細胞凋亡;高表達多種抗凋亡基因,如bcl-2基因、核轉錄因子κB(NF-κB)基因、突變型p53基因及c-myc基因等。CD44+/CD24-乳腺癌CSC對放射療法不敏感(Phillips et al.,2006,Journal of the National Cancer Institute,98:1777-1785);CD133+的膠質瘤干細胞具有很強的DNA損傷修復能力,對放射療法不敏感(Bao et al.,Glioma stem cells promote radioresistance by preferential activation of the DNA damage response.,2006,nature,444:756-760);Dylla等也報道結腸癌CSC在化療后的裸鼠異種腫瘤模型中細胞比率增高,對化療不敏感(Dylla et al.,Colorectal cancer stem cells are enriched in xenogeneic tumors following chemotherapy.,2008,PloS one,3:e2428)。且腫瘤干細胞和正常干細胞都高表達ATP結合盒轉運蛋白(ATP-binding cassette transporters,ABC),如ABCB1蛋白和ABCG2蛋白,前者編碼P-糖蛋白,后者是從米托蒽醌耐藥細胞中鑒定出來的。ABCB1和ABCG2介導的外排作用使Hoechst 33342和rhodamine 123等染料不能染色干細胞,證實了ABC蛋白是干細胞對化療不敏感特性的關鍵(Zhou et al.,The ABC transporter Bcrp1/ABCG2is expressed in a wide variety of stem cells and is a molecular determinant of the side-population phenotype.,2001,Nature medicine,7:1028-1034)。CSC具有和正常干細胞相似的多種生物學屬性,如相對增殖不活躍、對藥物和毒素不敏感、高表達多種ABC蛋白、核酸修復能力強、和凋亡抵抗能力(Dean et al.,Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell.,2005,Nature Reviews Cancer,5:275-284)。另外,CSC與正常干細胞類似,與周圍的微環境共同形成腫瘤干細胞龕,龕內相對缺氧且能向CSC提供大量的維持穩態的生物信號,對外界刺激具有屏障作用。常規的細胞毒性治療對處在細胞龕內的干細胞影響極小(LaBarge,The difficulty of targeting cancer stem cell niches.,2010,Clinical cancer research,16:3121-3129)。因此,CSC可以在放化療的 過程中存活下來,并且重新繁衍腫瘤,最終使得腫瘤復發和治療性耐受。
4)促轉移侵襲能力強。由于腫瘤組織的高度異質性,大部分腫瘤實體細胞并沒有侵襲及轉移的能力,而CSC則具有非常強的侵襲及轉移能力。如有報道顯示,乳腺癌CSC和乳腺癌轉移過程中重要的EMT過程高度相關,EMT樣的CSC往往定位在于實體腫瘤浸潤邊緣,并且很容易進入血液循環并在遠端形成微小轉移灶(Mani et al.,The epithelial-mesenchymal transition generates cells with properties of stem cells.,2008,Cell,133:704-715)。這時,CSC又可以在一些微環境因子(如ID1介導的TWIST下調(Stankic et al.,TGF-β-Id1signaling opposes Twist1and promotes metastatic colonization via a mesenchymal-to-epithelial transition.,2013,Cell reports,5:1228-1242))作用下轉變為高增值活性的上皮態CSC,最終完成乳腺癌轉移過程。在胰腺癌中,表達CD133+/CXCR4+細胞表面標記的的CSC也常常定位在腫瘤浸潤邊緣,容易進入血液,且其高表達趨化因子受體。在CXCR4趨化因子的作用下,該類CSC很容易轉移到肝臟中(Hermann et al.,Distinct populations of cancer stem cells determine tumor growth and metastatic activity in human pancreatic cancer.,2007,Cell stem cell,1:313-323)。上述研究均提示CSC在惡性腫瘤的浸潤和轉移中起著非常重要的作用。
綜上所述,CSC具有致瘤性強、復雜的異質性及生物可塑性、轉移侵襲能力強、且治療敏感性差。腫瘤組織經過常規的細胞毒性治療后,如放化療和生物靶向治療后,殘存的高致瘤性CSC開始增強增殖活性并開始擴張并驅動腫瘤組織生長,表現為臨床腫瘤復發。在上述理論和研究的基礎上,眾多研究學者認為,靶向CSC細胞群的治療策略將補充現有治療策略的不足,并能顯著的提高腫瘤治療水平(Schaue and McBride,Opportunities and challenges of radiotherapy for treating cancer.,2015,Nature Reviews Clinical Oncology,12:527-540;Takebe et al.,Targeting Notch,Hedgehog,and Wnt pathways in cancer stem cells:clinical update.,2015,Nature Reviews Clinical Oncology)。
目前的研究已明確,CSC中存在失調的信號通路,其對CSC的調控至關重要,包括Wnt信號通路,Notch信號通路,和Hedgehog信號通路等(Takebe et al.,Targeting Notch,Hedgehog,and Wnt pathways in cancer stem cells:clinical update.,2015,Nature Reviews Clinical Oncology)。1916年,Notch基因首次在果蠅體內被發現,該基因的突變體可使果蠅翅膀邊緣產生缺口,因此得名。完整的Notch信號通路包括跨膜蛋白受體、跨膜蛋白、轉錄因子及下游靶基因,其中人類細胞共有4種Notch受體(Notch1~4)及5種配體(Delta-like-1,Delta-like-3,Delta-like-4,Jagged-1和Jagged-2)(Bailey et al.,Cancer metastasis facilitated by developmental pathways:Sonic hedgehog,Notch,and bone morphogenic proteins.,2007,Journal of cellular biochemistry,102:829-839),其在進化中高度保守,具有調控細胞增殖、分化和凋亡的功能,并幾乎涉及所有組織和器官。Notch基因具有十分顯著的致瘤性作用,這體現在多種人類腫瘤(包括淋巴瘤、乳腺癌、肺癌、頭頸腫瘤、胰腺癌、結腸癌、頜骨骨肉瘤和膠質母細胞瘤)的激活突變,并擴大到Notch信號通路的各級成員(Weng and Aster,Multiple niches for Notch in cancer:context is everything.,2004,Current opinion in genetics&development,14:48-54)。在多種腫瘤中,Notch信號通路被證實跟腫瘤干細胞干性的維持有關。最近,OncoMed制藥公司公布靶向Notch通路的單克隆抗體tarextumab治療小細胞肺癌的Ⅰb期臨床試驗的患者存活和生物標記資料。tarextumab是完全人源化單克隆抗體,靶向Notch2/3受體。臨床前研究提示有2種作用機制,即下調Notch通路信號,抗腫瘤干細胞作用和影響周細胞、基質細胞和腫瘤的微環境。
而在實體腫瘤細胞中,表皮生長因子受體(EGFR)信號是機體的另一條重要信號轉導途徑,廣泛存在于上皮、間質及神經組織,貫穿于整個細胞的生長和增殖過程,對細胞的分化選擇起重要的調節作用。EGFR家族主要包括EGFR(即HER1)和HER2-4。EGFR編碼跨膜蛋白,其信號經受體酪氨酸磷酸化被激活,通過Ras/Raf/MEK/MAPK途徑級聯放大,最后導致MAPK的磷酸化,修飾的MAPK信號進入細胞核,促進目標基因的磷酸化,調節基因的表達和活性。EGFR信號與腫瘤的發生發展及預后密切相關,在多種腫瘤如肺癌、結腸癌、乳腺癌、前列腺癌、卵巢癌及膀膚癌等中均有表達。高表達的信號與乳腺癌和膀朧癌的不良預后相關。EGFR在乳腺癌中的表達與腫瘤增殖,疾病進展,以及不良預后相關一。的表達可降低雌激素受體的表達,增加對內分泌治療的抵抗,。臨床前研究表明,組成性活化的EGFR可明顯增加體內MCF-9人乳癌模型的成瘤性。作用于表皮生長因子受體(epithelial growth factor receptor,EGFR)的單克隆抗體西妥昔單抗(cetuximab,C225)是研究最為廣泛的靶向藥物之一,其療效在多種腫瘤治療中得到肯定。
EGFR與Notch信號的相互作用方式復雜,可以表現為協同作用,也可以表現為拮抗作用,依賴于組織的類型和發育的階段。在神經膠質瘤中,Notch上調EGFR的表達,在高分化神經膠質瘤中,EGFR的表達與Notch密切相關。而在皮膚鱗狀細胞癌中,EGFR信號作用于Notch1信號的上游,并且抑制信號的表達。與信號在乳腺原位導管細胞癌的體外培養中發揮協同促進作用。這些研究表明EGFR信號與Notch信號是緊密聯系的兩條信號傳導通路,對于腫瘤的發生發展具有重要的調節作用。
綜上所述,Notch信號通路是參與腫瘤細胞EMT的發生、腫瘤CSC細胞群及干性的維持及其增殖分化、以及CSC治療耐受性調控中的關鍵通路。而EGFR是腫瘤實體細胞中主要 的一類過度激活信號通路,且在腫瘤實體瘤靶向治療中占有重要地位。而兩個信號通路間存在各個層次上的交叉激活。
因此,采用基因工程技術構建一個可以同時阻斷兩種信號的多特異性抗體,既能阻斷EGFR,又能阻斷Notch,進而同時對腫瘤干細胞及腫瘤實體細胞產生抑制作用一直是本領域的技術人員急待解決的問題。
技術實現要素:
本發明的目的在于提供了一個可以同時阻斷兩種信號通路的多特異性抗體,即可以阻斷EGFR,也能阻斷Notch2和Notch3。本發明的另一目的在于提供該抗體的制備方法。本發明的第三目的在于利用該抗體,以及包含該抗體作為活性成分的用于腫瘤疾病的治療劑。
本發明的第一方面,提供一種抗EGFR和Notch2/3IgG分子樣交叉單克隆抗體(Crossmab)C-T Crossmab,該Crossmab能與EGFR,Notch2,及Notch3結合。
優選的,本發明的一種抗人EGFR和Notch多特異性抗體,其包括兩條重鏈多肽和兩條輕鏈多肽,其中重鏈多肽的氨基酸序列如SEQ ID NO:1和SEQ ID NO:2所示;輕鏈多肽的氨基酸序列如SEQ ID NO:3和SEQ ID NO:4所示。
本發明還提供了一種多核苷酸,其包含編碼如上所述的抗體的核苷酸序列。
優選的,所述的多核苷酸,其包含編碼重鏈多肽的如SEQ ID NO:5和SEQ ID NO:6所示的核苷酸序列;以及編碼輕鏈多肽的如SEQ ID NO:7和SEQ ID NO:8所示的核苷酸序列。
本發明還提供了一種重組表達載體,其包括如上所述的多核苷酸。
優選的,所述的重組表達載體,選自pcDNA3.1、pDR1或pDHFR等。在本發明的優選實施例中,表達載體具體為pcDNA3.1。
本發明還提供了一種用上述的載體轉化的宿主細胞。
優選的,所述的宿主細胞,為真核細胞;更優的,選自中國倉鼠卵巢CHO細胞、NS0骨髓瘤、COS或SP2/0細胞等。
在本發明中,任何編碼EGFR、Notch2和Notch3的合適的DNA都適合于本發明,這樣的結構包括從組織或細胞mRNA中分離得到的,從根據以公布數據庫中序列全基因合成的,或從其他cDNA文庫中獲得的。
在本發明中,任何合適的載體都可以使用,可以為pDR1,pCDNA3.1,pDHFF之一,表達載體中包括連接有合適的轉錄和翻譯調節序列的融合DNA序列。
哺乳動物或昆蟲宿主細胞培養系統可用于本發明的C-T Crossmab的表達,COS、CHO、NS0、sf9及sf21等均可適用于本發明。
可用的宿主細胞為含有上述載體的原核細胞,可以為DH5a、BL21(DE3)、TG1之一。
本發明的第二方面,提供了上述抗體(C-T Crossmab)的制備方法,其包括如下步驟:
(i)在適合允許所述抗體表達的條件下,培養根據如前所述的宿主細胞;和
(ii)回收表達的抗體。
步驟(ii),進一步分離或純化回收表達的抗體。
本發明中公開的C-T Crossmab的制備方法為在表達條件下,培養上述的宿主細胞,從而表達C-T Crossmab,分離或純化所述的C-T Crossmab。
利用上述方法,可以將雙特異性抗體純化為基本均一的物質,例如在SDS-PAGE電泳上為單一條帶。
可以利用親和層析的方法對本發明公開的C-T Crossmab進行分離純化,根據所利用的親和柱的特性,可以使用常規的方法例如高鹽緩沖液、改變PH等方法洗脫結合在親和柱上的C-T Crossmab多肽。
在本發明的一個優選實施例中,本發明公開的上述多特異性抗體C-T Crossmab是通過以下方法得到的:全基因合成多特異性抗體的重輕鏈基因,全基因的序列參考了單克隆抗體cetuximab的序列(美國專利US6217866B1,Li et al.,Structural basis for inhibition of the epidermal growth factor receptor by cetuximab.,2005,Cancer Cell,7:301-311)、tarextumab的序列(美國專利US20130323266 A1,Yen,Wan-Ching,et al..,題名Targeting Notch signaling with a Notch2/Notch3antagonist(tarextumab)inhibits tumor growth and decreases tumor-initiating cell frequency.,linical Cancer Research 21.9(2015):2084-2095)及Crossmab重鏈輕鏈設計方案(Hu et al.,Four-in-one antibodies have superior cancer inhibitory activity against EGFR,HER2,HER3,and VEGF through disruption of HER/MET crosstalk.,2015,Cancer research,75:159-170)和(Spasevska et al.,Advances in Bispecific Antibodies Engineering:Novel Concepts for Immunotherapies.,2015,J Blood Disorders Transf,6:2)。
本發明是用全基因合成多特異性抗體的重鏈基因和輕鏈基因,并將上述兩條重鏈基因、兩條輕鏈基因分別裝入真核表達載體pcDNA3.1和pcDNA3.1Zeo。上述質粒一起用脂質體法轉染CHO-K1細胞,并用含500μg/ml G418和300μg/ml Zeocin的選擇培養基篩選穩定表達雙特異性抗體的細胞克隆.利用Protein A柱通過親和層析從細胞培養物的上清中純化多特異性抗體C-T Crossmab。
本發明的第三方面,提供了所述的抗體(C-T Crossmab)在制備抗腫瘤藥物中的應用。
所述的藥物,以本發明的C-T Crossmab為活性成份。
本發明還提供了一種藥物組合物,其包含C-T Crossmab,和至少一種可藥用的載體、稀釋劑或賦形劑。
所述的藥物組合物在制備抗腫瘤藥物中的用途。
以上的用途,還包括和其它的抗腫瘤藥物聯合使用。
本發明對上述的C-T Crossmab進行親和力檢測,發現C-T Crossmab同時完好地保留了cetuximab和tarextumab的親和力。
因此,我們進行下一步實驗,包括抑制腫瘤細胞增殖、體內抑瘤和抑制腫瘤EMT基因表達等實驗,實驗結果表明,本發明公開的C-T Crossmab同時具有Cetuximab和tarextumab的功能。此外,該多特異性抗體C-T Crossmab可以和傳統的IgG分子一樣,最大程度地保留了傳統單克隆抗體結構,因為有Fc片段的存在,可以用普通的ProteinA柱親和層析法純化,利于大規模的生產純化。實驗表明,在相等劑量下,多特異性抗體C-T Crossmab具有與聯合應用cetuximab和tarextumab相似或更強的抗腫瘤治療效果。
本發明公開上述雙特異性抗體C-T Crossmab,可以和藥學上可以接受的輔料一起組成藥物制劑組合物從而更穩定地發揮療效,這些制劑可以保證本發明公開的雙特異性抗體氨基酸核心序列的構像完整性,同時還要保護蛋白質的多官能團防止其降解(包括但不限于凝聚、脫氨或氧化)。通常情況下,對于液體制劑,通常可以在2℃-8℃條件下保存至少穩定一年,對于凍干制劑,在30℃至少六個月保持穩定。在這里制劑可為制藥領域常用的混懸、水針、凍干等制劑,優選水針或凍干制劑,對于本發明公開的上述C-T Crossmab的水針或凍干制劑,藥學上可以接受的輔料包括表面活性劑、溶液穩定劑、等滲調節劑和緩沖液之一或其組合,其中表面活性劑包括非離子型表面活性劑如聚氧乙烯山梨醇脂肪酸酯(吐溫20或80);poloxamer(如poloxamer 188);Triton;十二烷基硫酸鈉(SDS);月桂硫酸鈉;十四烷基、亞油基或十八烷基肌氨酸;Pluronics;MONAQUATTM等,其加入量應使雙功能雙特異性抗體蛋白的顆粒化趨勢最小,溶液穩定劑可以為糖類,包括還原性糖和非還原性糖,氨基酸類包括谷氨酸單鈉或組氨酸,醇類包括三元醇、高級糖醇、丙二醇、聚乙二醇之一或其組合,溶液穩定劑的加入量應該使最后形成的制劑在本領域的技術人員認為達到穩定的時間內保持穩定狀態,等滲調節劑可以為氯化鈉、甘露醇之一,緩沖液可以為TRIS、組氨酸緩沖液、磷酸鹽緩沖液之一。
上述制劑為包含C-T Crossmab的組合物,在對包括人在內的動物給藥后,抗腫瘤效果明顯。具體來講,對腫瘤的預防和/或治療有效,可以作為抗腫瘤藥物使用。
本發明所指的腫瘤,包括腺癌、白血病、淋巴瘤、黑色素瘤、肉瘤,腫瘤組織的來源包 括但不限于腎上腺、膽囊、骨、骨髓、腦、乳腺、膽管、胃腸道、心臟、腎臟、肝臟、肺、肌肉、卵巢、胰腺、甲狀旁腺、陰莖、前列腺、皮膚、唾液腺、脾臟、睪丸、胸腺、甲狀腺和子宮。除了上述的腫瘤外,還可用于中樞神經系統的腫瘤如膠質細胞多樣性瘤、星細胞瘤等,此外眼部的腫瘤包括基底細胞癌、鱗狀細胞癌、黑色素瘤等,還包括內分泌腺腫瘤、神經內分泌系統腫瘤、胃腸道胰腺內分泌系統腫瘤,生殖系統腫瘤及頭頸部腫瘤等。這里不再一一列舉。
本發明所稱的抗腫瘤藥物,指具有抑制和/或治療腫瘤的藥物,可以包括伴隨腫瘤生長相關癥狀發展的延遲和/或這些癥狀嚴重程度的降低,它進一步還包括已存在的腫瘤生長伴隨癥狀的減輕并防止其他癥狀的出現,還也減少或防止轉移。
本發明中C-T Crossmab及其組合物在對包括人在內的動物給藥時,給藥劑量因病人的年齡和體重,疾病特性和嚴重性,以及給藥途徑而異,可以參考動物實驗的結果和種種情況,總給藥量不能超過一定范圍。具體講靜脈注射的劑量是1~1800mg/天。
本發明公開的C-T Crossmab及其組合物還可以和其他的抗腫瘤藥聯合給藥或放射治療,用于腫瘤的治療,這些抗腫瘤藥包括1、細胞毒類藥物(1)作用于DNA化學結構的藥物:烷化劑如氮芥類、亞硝尿類、甲基磺酸酯類;鉑類化合物如順鉑、卡鉑和草酸鉑等;絲裂霉素(MMC);(2)影響核酸合成的藥物:二氫葉酸還原酶抑制劑如甲氨喋呤(MTX)和Alimta等;胸腺核苷合成酶抑制劑如氟尿嘧啶類(5FU、FT-207、卡培他濱)等;嘌呤核苷合成酶抑制劑如6-巰基嘌呤(6-MP)和6-TG等;核苷酸還原酶抑制劑如羥基脲(HU)等;DNA多聚酶抑制劑如阿糖胞苷(Ara-C)和健擇(Gemz)等;(3)作用于核酸轉錄的藥物:選擇性作用于DNA模板,抑制DNA依賴RNA聚合酶,從而抑制RNA合成的藥物如:放線菌素D、柔紅霉素、阿霉素、表阿霉素、阿克拉霉素、光輝霉素等;(4)主要作用于微管蛋白合成的藥物:紫杉醇、泰索帝、長春花堿、長春瑞濱、鬼臼鹼類、高三尖杉酯堿;(5)其他細胞毒藥:門冬酰胺酶主要抑制蛋白質的合成;2、激素類抗雌激素:三苯氧胺、屈洛昔芬、依西美坦等;芳香化酶抑制劑:氨魯米特、蘭特隆、來曲唑、瑞寧德等;抗雄激素:氟它氨RH-LH激動劑/拮抗劑:諾雷德、依那通等;3、生物反應調節劑:主要通過機體免疫功能抑制腫瘤干擾素;白細胞介素-2;胸腺肽類;4、單克隆抗體:美羅華(MabThera);赫賽汀(Trastuzumab)Bevacizumab(Avastin);5、各種放射療法;6、其他括一些目前機制不明和有待進一步研究的藥物;細胞分化誘導劑如維甲類;細胞凋亡誘導劑。本發明公開的C-T Crossmab及其組合物可以和上述的抗腫瘤藥物之一或其組合聯合用藥。
本發明提供了一個可以同時阻斷EGFR,和Notch2和Notch3兩種信號通路的多特異性 抗體,本發明的多特異性抗體可用于腫瘤疾病的治療劑。
附圖說明
圖1.本發明C-T Crossmab結構示意圖;
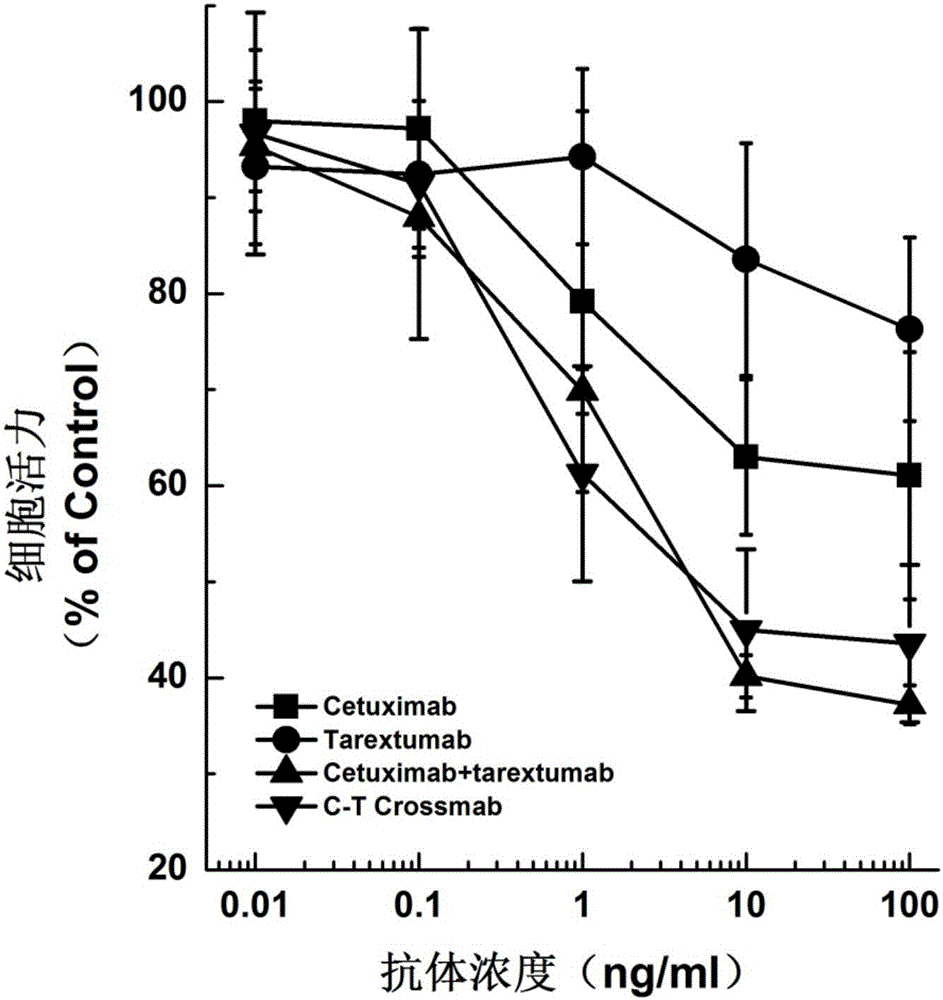
圖2.本發明C-T Crossmab抑制HCC827細胞活性實驗結果;
圖3.本發明C-T Crossmab抑制HCC827腫瘤生成曲線。
具體實施方式
下面結合實施例、實驗例和附圖對本發明進行進一步的說明,不應理解為對本發明的限制。實施例不包括對傳統方法的詳細描述,如那些用于構建載體和質拉的方法,將編碼蛋白的基因插入到這樣的載體和質拉的方法或將質粒引入宿主細胞的方法.這樣的方法對于本領域中具有普通技術的人員是眾所周知的,并且在許多出版物中都有所描述,包括Sambrook,J.,Fritsch,E.F.and Maniais,T.(1989)Molecular Cloning:A Laboratory Manual,2ndedition,Cold spring Harbor Laboratory Press
實施例1.本發明多功能抗體C-T Crossmab的構建及表達
全基因合成C-T Crossmab的兩條重鏈及兩條輕鏈基因序列(序列如SEQ ID NO:5、6、7和8所示,委托金唯智生物公司合成),兩條重鏈基因序列與pcDNA3.1載體相連,兩條輕鏈基因序列與pcDNA3.1Zeo載體相連,構建成真核表達載體。
于3.5cm組織培養皿中接種3×105CHO-K1細胞,細胞培養至80%-85%融合時進行轉染:取重鏈質粒各10μg,輕鏈質粒各4μg和30μl Lipofectamine2000 Reagent(Invitrogen公司產品)分別溶于800μl無血清DMEM培養基,室溫靜置5分鐘,將以上2種液體混合,室溫孵育20分鐘以使DNA-脂質體復合物形成,其間用3ml無血清的DMEM培養基替換培養皿中的含血清培養基,然后將形成的DNA-脂質體復合物加入到板中,CO2孵箱培養4小時后補加2ml含10%血清的DMEM完全培養基,置于CO2孵箱中繼續培養。轉染進行24h后細胞換含500μg/ml G418和300μg/ml Zeocin的選擇培養基篩選抗性克隆。將篩選得到的高表達克隆用無血清培養基擴大培養,用Protein A親和柱(GE公司產品)分離純化雙特異性抗體C-T Crossmab。將C-T Crossmab用PBS進行透析,最后以紫外吸收法定量。C-T Crossmab的結構如圖1所示,氨基酸序列如SEQ ID NO:1、2、3、4所示,委托金唯智生物公司測序成功。
實施例2.Biacore分析
將多克隆抗人FC抗體(Jackson ImmunoResearch公司)包被在CM5M5芯片(GE公司) 上,捕獲被檢測抗體后,用Biacore T100(GE Healthcare)檢測C-T Crossmab的親和力,具體檢測親和力數值見表1。
表1.Biacore分析
實驗結果說明,本發明的C-T Crossmab同時完好地保留了cetuximab和tarextumab的親和力。
實施例3:C-T Crossmab抑制非小細胞肺癌細胞活力實驗
取生長狀態良好的HCC827細胞(ATCC),調整細胞濃度為5×103/ml,接種于96孔細胞培養板,200μl/孔,于37℃、5%C02孵箱中培養24h后,在培養液中加入終濃度為5nmol的EGF和不同濃度梯度的雙特異性抗體C-T Crossmab,cetuximab,tarextumab,cetuximab+tarextumab,無關human IgG(Rituximab購自Roche公司),4天后,細胞活力用CellTiter-Glo Luminescent Cell Viability Assay試劑盒(Promega,Madison,WI)檢測。
實驗結果如圖2所示。實驗結果表明,C-T Crossmab可以抑制的HCC827細胞活力,比cetuximab和tarextumab有更顯著的抑制效果,與cetuximab及tarextumab聯合應用的雙抗體組效果相似或更優。
實施例4:C-T Crossmab體內抑制腫瘤生長實驗
為檢測C-T Crossmab體內抑瘤活性,首先用HCC827細胞,接種于到BALB/c裸鼠(第二軍醫大學實驗動物中心)右脅側皮下,成瘤后尾靜脈注射10mg/kg各組下列抗體:C-T Crossmab,cetuximab,tarextumab,無關對照human IgG,cetuximab和tarextumab兩種單抗各5mg/kg聯合應用,每周注射1次,持續至小鼠腫瘤過大處死。每天測量腫瘤的長寬,計算腫瘤體積。
腫瘤生長曲線如圖3所示。實驗結果表明:C-T Crossmab治療組腫瘤生長速度顯著小于cetuximab及tarextumab治療組(70天后,P<0.01,Mann-Whitney檢驗)。且C-T Crossmab的治療效果優于與cetuximab和tarextumab聯合應用效果(70天后,P<0.01,Mann-Whitney檢驗)。
以上已對本發明創造的較佳實施例進行了具體說明,但本發明創造并不限于所述實施例,熟悉本領域的技術人員在不違背本發明創造精神的前提下還可做出種種的等同的變型或替換,這些等同的變型或替換均包含在本申請權利要求所限定的范圍內。
SEQUENCE LISTING
<110> 中國人民解放軍第二軍醫大學
<120> 抗人EGFR和Notch多特異性抗體、其制備方法及用途
<130> 說明書,權利要求書
<160> 8
<170> PatentIn version 3.3
<210> 1
<211> 449
<212> PRT
<213> 人工序列
<400> 1
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asn Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr Asn Thr Pro Phe Thr
50 55 60
Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ser Asn Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 2
<211> 449
<212> PRT
<213> 人工序列
<400> 2
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Ser
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Ala Ser Ser Gly Ser Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ile Phe Tyr Thr Thr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro
115 120 125
Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu
130 135 140
Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn
145 150 155 160
Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser
165 170 175
Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala
180 185 190
Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly
195 200 205
Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Ser
355 360 365
Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 3
<211> 214
<212> PRT
<213> 人工序列
<400> 3
Asp Ile Leu Leu Thr Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser
65 70 75 80
Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 4
<211> 213
<212> PRT
<213> 人工序列
<400> 4
Asp Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Arg Ser Asn
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Ser Asn Phe Pro
85 90 95
Ile Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser
100 105 110
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr
115 120 125
Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
130 135 140
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
145 150 155 160
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
165 170 175
Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr Tyr Thr
180 185 190
Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Thr Val
195 200 205
Glu Arg Lys Cys Cys
210
<210> 5
<211> 1347
<212> DNA
<213> 人工序列
<400> 5
caggtgcagc tgaagcagag cggccccggc ctggtgcagc ccagccagag cctgagcatc 60
acctgcaccg tgagcggctt cagcctgacc aactacggcg tgcactgggt gcgccagagc 120
cccggcaagg gcctggagtg gctgggcgtg atctggagcg gcggcaacac cgactacaac 180
acccccttca ccagccgcct gagcatcaac aaggacaaca gcaagagcca ggtgttcttc 240
aagatgaaca gcctgcagag caacgacacc gccatctact actgcgcccg cgccctgacc 300
tactacgact acgagttcgc ctactggggc cagggcaccc tggtgaccgt gagcgccgcc 360
agcaccaagg gccccagcgt gttccccctg gcccccagca gcaagagcac cagcggcggc 420
accgccgccc tgggctgcct ggtgaaggac tacttccccg agcccgtgac cgtgagctgg 480
aacagcggcg ccctgaccag cggcgtgcac accttccccg ccgtgctgca gagcagcggc 540
ctgtacagcc tgagcagcgt ggtgaccgtg cccagcagca gcctgggcac ccagacctac 600
atctgcaacg tgaaccacaa gcccagcaac accaaggtgg acaagcgcgt ggagcccaag 660
agctgcgaca agacccacac ctgccccccc tgccccgccc ccgagctgct gggcggcccc 720
agcgtgttcc tgttcccccc caagcccaag gacaccctga tgatcagccg cacccccgag 780
gtgacctgcg tggtggtgga cgtgagccac gaggaccccg aggtgaagtt caactggtac 840
gtggacggcg tggaggtgca caacgccaag accaagcccc gcgaggagca gtacaacagc 900
acctaccgcg tggtgagcgt gctgaccgtg ctgcaccagg actggctgaa cggcaaggag 960
tacaagtgca aggtgagcaa caaggccctg cccgccccca tcgagaagac catcagcaag 1020
gccaagggcc agccccgcga gccccaggtg tacaccctgc ccccctgccg cgaggagatg 1080
accaagaacc aggtgagcct gtggtgcctg gtgaagggct tctaccccag cgacatcgcc 1140
gtggagtggg agagcaacgg ccagcccgag aacaactaca agaccacccc ccccgtgctg 1200
gacagcgacg gcagcttctt cctgtacagc aagctgaccg tggacaagag ccgctggcag 1260
cagggcaacg tgttcagctg cagcgtgatg cacgaggccc tgcacaacca ctacacccag 1320
aagagcctga gcctgagccc cggcaag 1347
<210> 6
<211> 1347
<212> DNA
<213> 人工序列
<400> 6
gaggtgcagc tggtggagag cggcggcggc ctggtgcagc ccggcggcag cctgcgcctg 60
agctgcgccg ccagcggctt caccttcagc agcagcggca tgagctgggt gcgccaggcc 120
cccggcaagg gcctggagtg ggtgagcgtg atcgccagca gcggcagcaa cacctactac 180
gccgacagcg tgaagggccg cttcaccatc agccgcgaca acagcaagaa caccctgtac 240
ctgcagatga acagcctgcg cgccgaggac accgccgtgt actactgcgc ccgcagcatc 300
ttctacacca cctggggcca gggcaccctg gtgaccgtga gcagcgccag cgtggccgcc 360
cccagcgtgt tcatcttccc ccccagcgac gagcagctga agagcggcac cgccagcgtg 420
gtgtgcctgc tgaacaactt ctacccccgc gaggccaagg tgcagtggaa ggtggacaac 480
gccctgcaga gcggcaacag ccaggagagc gtgaccgagc aggacagcaa ggacagcacc 540
tacagcctga gcagcaccct gaccctgagc aaggccgact acgagaagca caaggtgtac 600
gcctgcgagg tgacccacca gggcctgagc agccccgtga ccaagagctt caaccgcggc 660
gagtgcgaca agacccacac ctgccccccc tgccccgccc ccgagctgct gggcggcccc 720
agcgtgttcc tgttcccccc caagcccaag gacaccctga tgatcagccg cacccccgag 780
gtgacctgcg tggtggtgga cgtgagccac gaggaccccg aggtgaagtt caactggtac 840
gtggacggcg tggaggtgca caacgccaag accaagcccc gcgaggagca gtacaacagc 900
acctaccgcg tggtgagcgt gctgaccgtg ctgcaccagg actggctgaa cggcaaggag 960
tacaagtgca aggtgagcaa caaggccctg cccgccccca tcgagaagac catcagcaag 1020
gccaagggcc agccccgcga gccccaggtg tgcaccctgc cccccagccg cgaggagatg 1080
accaagaacc aggtgagcct gagctgcgcc gtgaagggct tctaccccag cgacatcgcc 1140
gtggagtggg agagcaacgg ccagcccgag aacaactaca agaccacccc ccccgtgctg 1200
gacagcgacg gcagcttctt cctggtgagc aagctgaccg tggacaagag ccgctggcag 1260
cagggcaacg tgttcagctg cagcgtgatg cacgaggccc tgcacaacca ctacacccag 1320
aagagcctga gcctgagccc cggcaag 1347
<210> 7
<211> 642
<212> DNA
<213> 人工序列
<400> 7
gacatcctgc tgacccagag ccccgtgatc ctgagcgtga gccccggcga gcgcgtgagc 60
ttcagctgcc gcgccagcca gagcatcggc accaacatcc actggtacca gcagcgcacc 120
aacggcagcc cccgcctgct gatcaagtac gccagcgaga gcatcagcgg catccccagc 180
cgcttcagcg gcagcggcag cggcaccgac ttcaccctga gcatcaacag cgtggagagc 240
gaggacatcg ccgactacta ctgccagcag aacaacaact ggcccaccac cttcggcgcc 300
ggcaccaagc tggagctgaa gcgcaccgtg gccgccccca gcgtgttcat cttccccccc 360
agcgacgagc agctgaagag cggcaccgcc agcgtggtgt gcctgctgaa caacttctac 420
ccccgcgagg ccaaggtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 540
ctgagcaagg ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 600
ctgagcagcc ccgtgaccaa gagcttcaac cgcggcgagt gc 642
<210> 8
<211> 639
<212> DNA
<213> 人工序列
<400> 8
gacatcgtgc tgacccagag ccccgccacc ctgagcctga gccccggcga gcgcgccacc 60
ctgagctgcc gcgccagcca gagcgtgcgc agcaactacc tggcctggta ccagcagaag 120
cccggccagg ccccccgcct gctgatctac ggcgccagca gccgcgccac cggcgtgccc 180
gcccgcttca gcggcagcgg cagcggcacc gacttcaccc tgaccatcag cagcctggag 240
cccgaggact tcgccgtgta ctactgccag cagtacagca acttccccat caccttcggc 300
cagggcacca aggtggagat caagagcagc gccagcacca agggccccag cgtgttcccc 360
ctggccccct gcagccgcag caccagcgag agcaccgccg ccctgggctg cctggtgaag 420
gactacttcc ccgagcccgt gaccgtgagc tggaacagcg gcgccctgac cagcggcgtg 480
cacaccttcc ccgccgtgct gcagagcagc ggcctgtaca gcctgagcag cgtggtgacc 540
gtgcccagca gcaacttcgg cacccagacc tacacctgca acgtggacca caagcccagc 600
aacaccaagg tggacaagac cgtggagcgc aagtgctgc 639
- 還沒有人留言評論。精彩留言會獲得點贊!
- 用于治療HBV感染和相關病癥的三特異性結合分子的制造方法與工藝
- 對磺胺類藥物具有特異性的固相微萃取纖維的制備方法與流程
- 一種檢測豬初乳中抗PEDV特異性IgA抗體的夾心ELISA試劑盒的制造方法與工藝
- 用于施用CD19xCD3雙特異性抗體的給藥方案的制造方法與工藝
- 雙特異性抗?VEGF/抗?ANG?2抗體及其在治療眼血管疾病中的應用的制造方法與工藝
- 特異性人EWS?FLI1融合蛋白抗體的制備方法及在制備尤文肉瘤診斷抗體試劑的應用與制造工藝
- 雙特異性四價抗體及其制造和使用方法與制造工藝
- 利用特異性揮發物篩選昆蟲相關嗅覺蛋白的方法與制造工藝
- 使用與表面偶聯的單特異性四聚抗體復合物組合物從樣品中分離出靶實體的方法與制造工藝
- 抗-WT1/HLA雙特異性抗體的制造方法與工藝
- 與CD56分子特異性結合的多肽及其應用的制造方法與工藝
- 用于治療HBV感染和相關病癥的三特異性結合分子的制造方法與工藝
- 對磺胺類藥物具有特異性的固相微萃取纖維的制備方法與流程
- 避免信號干擾的肌原纖維蛋白絲氨酸磷酸化水平的測定方法與流程
- 一種檢測豬初乳中抗PEDV特異性IgA抗體的夾心ELISA試劑盒的制造方法與工藝
- 用于施用CD19xCD3雙特異性抗體的給藥方案的制造方法與工藝
- 雙特異性抗?VEGF/抗?ANG?2抗體及其在治療眼血管疾病中的應用的制造方法與工藝
- 特異性人EWS?FLI1融合蛋白抗體的制備方法及在制備尤文肉瘤診斷抗體試劑的應用與制造工藝
- 活化人CD40的抗體和針對人PD?L1的抗體的組合療法的制造方法與工藝
- 雙特異性四價抗體及其制造和使用方法與制造工藝