利用乙醇醛合成乙酰輔酶A及其衍生產品的新途徑的制作方法
本發明涉及生物醫藥技術領域,具體涉及一種利用乙醇醛合成乙酰輔酶A及其衍生產品的方法。
背景技術:
乙酰輔酶A(AcCoA)是合成必要的生物化合物的關鍵中間體,該必要生物化合物包括聚酮化合物、脂肪酸、類異戊二烯、生物堿、維生素和氨基酸等。來源于AcCoA的代謝產物為初級和次級代謝物,其包括工業效用的化合物。AcCoA可以由乙酰磷酸(AcP)生成(如可通過磷酸乙酰轉移酶Pta(EC 2.3.1.8)催化得到),繼而生成一系列以AcCoA為平臺的生物產品,廣泛應用于生物催化等領域。AcP的合成方法有:
專利申請WO2015/144447A1公開了一種利用磷酸轉酮酶(EC 4.1.2.9、果糖-6-磷酸磷酸轉酮酶EC4.1.2.22)或磺基乙醛乙酰轉移酶(EC 2.3.3.15)催化甲醛生產乙酰磷酸的方法,反應方程如下:
2CH2O+磷酸→乙酰磷酸+H2O。
上述制備反應使用的磷酸轉酮酶,與甲醛的親和力差,催化效率不及轉化其最適底物5-磷酸木酮糖時的百分之一,并且微生物細胞對于甲醛的耐受能力普遍較差,可應用拓展的空間較小。
專利申請WO2015/181074A1公開了利用磷酸轉酮酶(EC 4.1.2.9、果糖-6-磷酸磷酸轉酮酶EC4.1.2.22)催化D-果糖生產D-赤蘚糖和乙酰磷酸的方法,該方法還包括將D-赤蘚糖轉化為乙醇醛,然后再進一步轉化為乙酰磷酸,反應過程如下:
上述乙酰磷酸的制備方法的反應速率較低,磷酸轉酮酶對于果糖的轉化數Kcat僅為0.1/s,效率很低,并且反應時間較長(18h)會對酶和產物的穩定性提出考驗。另外,第三步D-erythrose裂解為乙醇醛的反應是可逆過程,其反應速率受到裂解后的乙醇醛濃度的影響,使體系中很難積累高濃度的乙醇醛,這會導致乙酰磷酸的生成速率難以保證。第二步酶促反應生成的乙酰磷酸很可能會由于產物抑制而影響最后一步酶促反應的收率,如果第二步酶促反應生成乙酰磷酸后即時除去,會使整個制備工藝操作復雜化。
技術實現要素:
為克服現有技術的不足,本發明一方面提供一種5-磷酸阿拉伯糖(Ara5P)的制備方法,所述方法包括在酶催化下乙醇醛和3-磷酸甘油醛反應生成Ara5P,所述的酶選自:醛縮酶、轉醛醇酶、及其同工酶、突變酶。
優選的,所述的制備的反應是通過微生物來進行的;
優選的,所述的醛縮酶為果糖二磷酸醛縮酶(EC 4.1.2.13);
優選的,所述的轉醛醇酶為大腸桿菌Escherichia coli來源的轉醛醇酶(EC 2.2.1.2)的突變酶Tal BF178Y。
本發明另一方面還提供一種用于制備Ara5P的組合物,包括:
(1)乙醇醛,
(2)3-磷酸甘油醛,
(3)醛縮酶、轉醛醇酶、或其同工酶、突變酶;
或,
(1)乙醇醛,
(2)3-磷酸甘油醛,
(3)表達醛縮酶、轉醛醇酶、或其同工酶、突變酶的微生物;
優選的,所述的醛縮酶為果糖二磷酸醛縮酶(EC 4.1.2.13);
優選的,所述的轉醛醇酶為大腸桿菌Escherichia coli來源的轉醛醇酶(EC 2.2.1.2)的突變酶Tal BF178Y。
本發明另一方面提供一種乙酰磷酸(AcP)的制備方法,所述方法包括上述Ara5P的制備方法步驟;優選的,所述的AcP的制備方法還包括Ara5P轉化為5-磷酸木酮糖(Xu5P)再進一步轉化為AcP的步驟;
優選的,所述的Ara5P轉化為Xu5P的步驟包括Ara5P轉化為5-磷酸核酮糖(Ru5P)再進一步轉化為Xu5P的步驟;
優選的,所述的Xu5P轉化為AcP的反應包括Xu5P和磷酸在磷酸轉酮酶催化下生成AcP的步驟;
更優選的,所述的磷酸轉酮酶為磷酸轉酮酶(EC 4.1.2.9)或果糖-6-磷酸磷酸轉酮酶(EC 4.1.2.22);
所述的磷酸轉酮酶可來自各種種屬的微生物表達、人工合成及純化過程,特別是來源于微生物如優選來源于Pseudomonas stutzeri(如施氏假單胞菌Pseudomonas stutzeri A1501)和Bifidobacterium adolescentis菌株的磷酸轉酮酶在上述反應中均可發揮良好的催化作用;
優選的,Xu5P轉化為AcP的反應體系中還包括磷酸轉酮酶的輔酶,如焦磷酸硫胺素;
優選的,所述的制備的反應是通過微生物來進行的。
本發明另一方面提供一種用于制備AcP的組合物,包括上述用于制備Ara5P的組合物;
優選的,上述用于制備AcP的組合物還包括磷酸、磷酸轉酮酶;
更優選的,上述用于制備AcP的組合物還包括磷酸轉酮酶輔酶;
優選的,上述用于制備AcP的組合物還包括可催化Ara5P生成Xu5P的酶,該酶可為一種酶,也可為兩種以上的酶的組合,優選為核糖-5-磷酸異構酶和核酮糖-磷酸3-異構酶的組合;
在本發明的一個優選實施例中,上述用于制備AcP的組合物包括:
(1)乙醇醛,
(2)3-磷酸甘油醛,
(3)醛縮酶、轉醛醇酶、或其同工酶、突變酶,
(4)核糖-5-磷酸異構酶,
(5)核酮糖-磷酸3-異構酶,
(6)磷酸、磷酸轉酮酶、輔酶。
優選的,所述的醛縮酶為果糖二磷酸醛縮酶(EC 4.1.2.13);
優選的,所述的轉醛醇酶為大腸桿菌Escherichia coli來源的轉醛醇酶(EC 2.2.1.2)的突變酶Tal BF178Y;
優選的,所述的磷酸轉酮酶為磷酸轉酮酶(EC 4.1.2.9)或果糖-6-磷酸磷酸轉酮酶(EC 4.1.2.22);
優選的,所述的磷酸轉酮酶輔酶為焦磷酸硫胺素;
在本發明的一個更優選實施例中,上述用于制備AcP的組合物包括:
(1)乙醇醛,
(2)3-磷酸甘油醛,
(3)果糖二磷酸醛縮酶(EC 4.1.2.13)或大腸桿菌Escherichia coli來源的轉醛醇酶(Tal B)的突變酶Tal BF178Y,
(4)核糖-5-磷酸異構酶(EC 5.3.1.6),
(5)核酮糖-磷酸3-異構酶(EC 5.1.3.1),
(6)磷酸、磷酸轉酮酶(EC 4.1.2.9)或果糖-6-磷酸磷酸轉酮酶(EC4.1.2.22)及焦磷酸硫胺素。
AcP的穩定性較差,在實際應用中常做成乙酰磷酸鹽的形式,本發明另一方面提供一種乙酰磷酸鹽的制備方法,所述方法包括上述Ara5P的制備方法步驟;
優選的,所述的乙酰磷酸鹽的制備方法包括上述AcP的制備方法步驟;
優選的,所述的乙酰磷酸鹽包括:乙酰磷酸二鋰鹽、乙酰磷酸二鈉鹽、乙酰磷酸二銨鹽。
本發明另一方面提供一種乙酰輔酶A(AcCoA)的制備方法,所述方法包括上述Ara5P的制備方法步驟;優選的,所述的AcCoA的制備方法包括上述AcP的制備方法步驟;
優選的,所述的AcCoA的制備方法還包括在乙酰磷酸轉移酶(Pta)作用下AcP轉化為AcCoA的步驟;
優選的,所述的制備的反應是通過微生物來進行的。
本發明另一方面提供一種AcCoA衍生化合物的制備方法,所述方法包括上述Ara5P的制備方法步驟,優選的,上述AcP的制備方法步驟,更優選的,上述AcCoA的制備方法步驟;在本發明的一個優選實施例中,所述的AcCoA衍生化合物選自:丙酮、異丙醇、乙酸、L-谷氨酸、L-谷氨酰胺、L-脯氨酸、L-精氨酸、L-亮氨酸、L-半胱氨酸、琥珀酸酯和聚羥基脂肪酸酯;
在本發明的另一個優選實施例中,所述的AcCoA衍生化合物為聚3-羥基丁酸酯(PHB);
優選的,所述的制備的反應是通過微生物來進行的。
上述Ara5P、AcP、乙酰磷酸鹽、AcCoA、AcCoA衍生化合物的制備方法中所述的3-磷酸甘油醛可采用商購,也可采用現有技術中已知的方法制備得到,如利用葡萄糖、甘油、磷酸二羥基丙酮等制備得到,例如磷酸二羥基丙酮可以在丙糖磷酸異構酶作用下轉化為3-磷酸甘油醛;也可在上述制備方法的反應體系中加入3-磷酸甘油醛的制備反應物,實現在線制備;3-磷酸甘油醛的來源并不會限制本發明的范圍。
上述Ara5P、AcP、乙酰磷酸鹽、AcCoA、AcCoA衍生化合物的制備方法中所述的乙醇醛可由現有技術中已知的方法制備得到,如,乙醛的鹵代反應、糖類的裂解反應等,其中,利用甲醛制備乙醇醛的成本較低,優選的,上述乙醇醛是利用甲醛按照現有技術優選如“甲醛合成乙醇醛研究及其應用進展”中記載的乙醇醛的制備方法和工藝(辛坤,李青松,賈冰,等.甲醛合成乙醇醛研究及其應用進展.天然氣化工·C1化學與化工,2016(41):88-94)制備得到。
本發明另一方面提供一種上述醛縮酶、轉醛醇酶或其同工酶、突變酶在制備Ara5P、AcP、乙酰磷酸鹽、AcCoA、AcCoA衍生化合物中的應用。
本發明另一方面還提供一種表達上述醛縮酶、轉醛醇酶或其同工酶、突變酶的微生物在制備Ara5P、AcP、乙酰磷酸鹽、AcCoA、AcCoA衍生化合物中的應用。
本發明涉及的酶可來自各種微生物來源及人工改造獲得的同工酶、突變酶。
上述制備的反應通過微生物來進行,如反應中涉及的反應物中的一種或兩種以上,和/或,酶的一種或兩種以上可由微生物在線生產。
本發明提供的Ara5P的制備方法中醛縮酶或轉醛醇酶不依賴于輔酶,催化效率較高(Vmax大于50U/mg),且醛縮酶及轉醛醇酶的種類繁多,可優化空間很大。本發明中磷酸轉酮酶的催化底物為5-磷酸木酮糖,是其最適底物,親和力和酶活很高(Km約為10-45mM,Vmax為90-800U/mg),并且作為最后一步反應,該催化過程不可逆,使體系不受反應平衡的限制,既可以拉動醛縮酶和轉醛醇酶的速率,又可以最大限度地提高乙酰磷酸的收率及促進3-磷酸甘油醛的回補。提供的AcP的制備方法,催化速率較高,反應路線的碳理論得率為100%,沒有碳損失,G3P和酶均可循環使用,反應效率較高,成本有所降低,另外,來自磷酸戊糖途徑的輔助酶RpiA及Rpe,與底物親和力強、穩定性好、催化活性高。上述制備方法的該方法在補料發酵、連續發酵等可控制底物水平的生產過程中會發揮更為明顯的優勢。
附圖說明
圖1所示為本發明實施例1提供的生物合成途徑示意圖。
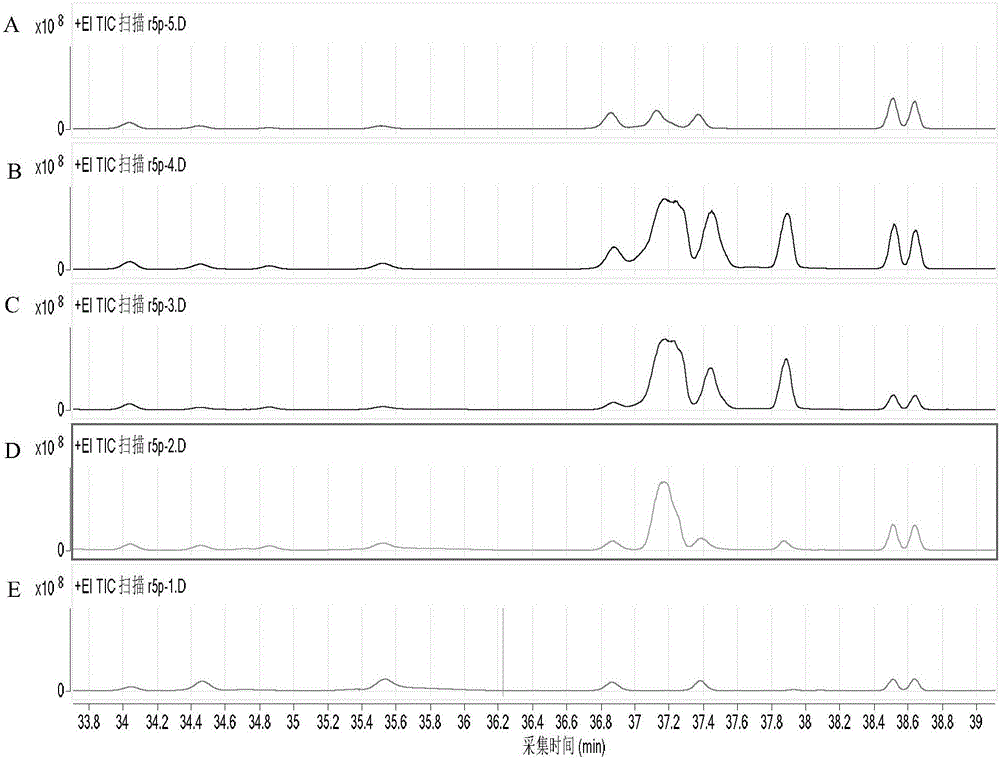
圖2所示為本發明實施例6提供的GC-MS檢測結果。
具體實施方式
下面將結合本發明實施例中的附圖,對本發明實施例中的技術方案進行清楚、完整地描述,顯然,所描述的實施例僅是本發明一部分實施例,而不是全部的實施例。基于本發明中的實施例,本領域普通技術人員在沒有作出創造性勞動前提下所獲得的所有其他實施例,都屬于本發明保護的范圍。
本發明的具體實施方式中涉及的利用乙醇醛生物合成乙酰磷酸的途徑如圖1所示,反應過程中涉及的反應物、中間體及酶的信息詳見表1和2。
表1本發明中涉及的代謝物信息表
表2本發明中涉及的酶信息表
實施例1
乙醇醛為15mM的初始濃度下,在大腸桿菌Escherichia coli來源的Transal dolase Tal B的突變酶Tal BF178Y(在原酶基礎上第178位氨基酸殘基由F突變為Y)作用下,添加5mM G3P,進行Ara5的制備反應,反應1h后,將反應液凍干、衍生處理后,通過GC-MS定量檢測到的Ara5P含量為2.5mM,Ara5P的平均合成速率為52.1μmol(Ara5P)/min/mg(酶蛋白)。
實施例2
乙醇醛在15mM的初始濃度下,ThDP濃度為1mM,PO43+濃度為2mM,加入Bifidobacterium adolescentis來源的Fpk(EC 4.1.2.22)、大腸桿菌來源的aldolase(Fsa,EC 4.1.2.13)、2mM G3P和微量的Rpi A、Rpe,進行AcP的制備反應。經過檢測,乙醇醛的轉化速率為16.3μmol(乙醇醛)/min/mg(酶蛋白)。
以上數據為反應體系進行3h的平均速率。
實施例3
乙醇醛在15mM的初始濃度下,ThDP濃度為1mM,PO43+濃度為2mM,加入施氏假單胞菌Pseudomonas stutzeri A1501來源的Xpk(EC 4.1.2.9)、大腸桿菌Escherichia coli來源的Transaldolase(Tal B,EC 2.2.1.2)的突變酶Tal BF178Y、2mM G3P和微量的Rpi A、Rpe,進行AcP的制備反應。經過檢測,乙醇醛的轉化速率為19.7μmol(乙醇醛)/min/mg(酶蛋白)。
以上數據為反應體系進行3h的平均速率。
實施例4
乙醇醛在20mM的初始濃度下,ThDP濃度為1mM,PO43+濃度為2mM,加入施氏假單胞菌Pseudomonas stutzeri A1501來源的Xpk、大腸桿菌Escherichiacoli來源的Transaldolase(tal B,EC 2.2.1.2)的突變酶Tal BF178Y、4mM DHAP和微量的Rpi A、Rpe、Tpi A,進行AcP的制備反應。經過檢測,乙醇醛的轉化速率為26.5μmol(乙醇醛)/min/mg(酶蛋白)。
以上數據為反應體系進行3h的平均速率。
實施例5
將合成磷酸轉酮酶的基因整合到大腸桿菌Escherichia coli K-12 MG1655的基因組中,然后將含有用于合成聚3-羥基丁酸酯(PHB)的基因(PhaA、PhaB和PhaC)以及Tal BF178Y的基因的質粒轉入到上述菌株中,上述質粒選擇I PTG誘導型啟動子(其余需要的酶,在大腸桿菌中均已存在)。將上述菌株在LB培養基中進行培養,培養2.5h后,菌體OD600值達到0.8-1.0時,添加IPTG至終濃度0.5mM,誘導(22℃,6h)表達上述質粒上的目標酶蛋白。離心(4℃、8000r/min、5min)收集菌體后,用不添加碳源和氮源的M9培養基懸浮菌體后,重新離心收集菌體,此懸浮、離心過程重復三次,用于除去菌體中殘留的碳源。將最終收集的菌體以上述M9培養基重新懸浮后,均分3份,分別添加0.02g、0.45g、0.9g乙醇醛(菌體中含有中間代謝物G3P,不需做額外添加),以上述M9培養基統一定容至30mL,發酵20h,觀察發酵過程中細胞生長情況,發酵結束后收集菌體并檢測發酵液中PHB含量。
PHB的檢測:將發酵液離心獲得菌體后,凍干稱重。利用體積比1:1的氯仿及酯化液(主要成分為甲醇)混合液4mL在100℃下對菌體凍干粉衍生4h,然后添加2mL超純水,靜置分層后,除去甲醇及細胞碎片等,取下層的3-羥基丁酸甲酯氯仿溶液,利用氣質聯用儀GC-MS對氯仿中的3-羥基丁酸甲酯進行定量檢測。
實驗結果:在上述發酵過程中,菌體可以進行正常的生長、繁殖;發酵結束后,相應的菌體干重分別為0.025g、0.076g、0.131g;經與標準品比對,測得樣品中3-羥基丁酸的總含量分別為0.005g、0.066g、0.089g,碳源轉化率在10%-20%之間。結果表明,菌體可以利用乙醇醛和自身含有的G3P、酶進行乙酰磷酸、乙酰輔酶A的合成,并繼續生成乙酰輔酶A的衍生物PHB用于自身的碳源儲備,乙醇醛在發酵過程中未表現出明顯的細胞毒性,并可維持菌體正常的代謝消耗。
實施例6
采用不同途徑制備乙酰磷酸和AcCoA,反應2h,反應結束后檢測Ara5P和AcCoA的含量,AcCoA的含量也可間接表示乙酰磷酸的合成量,反應物、酶及其添加量如表3所示。
Ara5P的檢測:取50μL反應液,將反應液凍干后,利用30μL甲氧胺鹽酸鹽和90μL三甲基硅基三氟乙酰胺分別衍生1h后,衍生溫度為37℃,利用GC-MS檢測Ara5P含量。
AcCoA的檢測:取50μL反應液,經10%的硫酸溶液終止反應后,用0.22μm的濾膜除去雜質后,進行液相檢測,流動相A為0.2M、pH=5的磷酸二氫鈉溶液,流動相B為乙腈,AcCoA的最大吸收峰出現在254nm。
產物含量檢測結果如表3所示。
表3結果表明,上述反應體系中不添加醛縮酶或轉醛醇酶時,沒有Ara5P生成,AcCoA的生成量也非常少,反應轉化率非常低;上述反應體系中添加醛縮酶或轉醛醇酶和G3P后,可生成Ara5P,且反應轉化率較高;在同時添加RpiA和Rpe后,生成的Ara5P會在RpiA、Rpe和F/Xpk作用下轉化為乙酰磷酸,進一步被磷酸乙酰轉移酶轉化為AcCoA,且由Ara5P和AcCoA的檢測結果可知,反應轉化率非常高;在體系中同時添加DHAP和TpiA,替代G3P,也可獲得與直接添加G3P相似的效果,達到較高的轉化率。
表3體系的物質添加和代謝物生成情況表
(反應物或產物濃度單位:mM;酶用量單位:μg)
備注:緩沖液中包含Tris、NaCl及MgCl2三種主要成分,pH7.5,37℃。
另外,在含有反應物如GA、Phos、G3P、ThDP等的相同反應體系中,分別:A:不添加醛縮酶或轉醛醇酶;B:添加Tal BF178Y;C:添加Tal BF178Y和F/Xpk;D:添加Tal BF178Y、F/Xpk以及Rpi A;E:添加Tal BF178Y、F/Xpk以及Rpi A和Rpe。GC-MS檢測Ara5P含量,結果如圖2中A-E圖所示(Ara5P的出峰時間為37min)。由圖2中各圖及其對應的反應條件可以看出,僅在添加醛縮酶后,Ara5P生成;僅在同時添加Rpi A和Rpe時,生成的Ara5P才會被磷酸轉酮酶較為徹底的分解。
以上所述僅為本發明的較佳實施例而已,并不用以限制本發明,凡在本發明的精神和原則之內,所作的任何修改、等同替換等,均應包含在本發明的保護范圍之內。
- 還沒有人留言評論。精彩留言會獲得點贊!
- 新的乙酰輔酶a羧化酶(acc)抑制劑及其在糖尿病、肥胖癥和代謝綜合征中的應用的制作方法
- 新的乙酰輔酶a羧化酶(acc)抑制劑及其在糖尿病、肥胖癥和代謝綜合征中的應用的制作方法
- 用作制備3
- 乙酰輔酶a羧化酶剪接變體及其用途的制作方法
- 新的乙酰輔酶a羧化酶(acc)抑制劑及其在糖尿病、肥胖癥和代謝綜合征中的應用的制作方法
- 氧肟酸類組蛋白去乙酰化酶抑制劑及其制備方法和用途的制作方法
- 人類病原體艱難梭菌中乙酰輔酶a合成酶的抑制劑的制作方法
- 作為乙酰輔酶a羧化酶(acc)抑制劑的螺苯并二氫吡喃-4-酮的制作方法
- 一種新的多肽——人過氧化物酶體脂肪酸乙酰輔酶a氧化酶46.97和編碼這種多肽的多核苷酸的制作方法
- 基于在線控制氧消耗速率的輔酶q10發酵新工藝的制作方法