GsHA12蛋白及其編碼基因在調控植物耐逆性中的應用的制作方法
本發明屬于生物技術領域,具體涉及gsha12蛋白及其編碼基因在調控植物耐逆性中的應用。
背景技術:
土地鹽堿化嚴重威脅我國乃至世界農業系統的正常發展,全世界約20%的可耕種地和50%的灌溉地受鹽堿化影響。中國東北地域的松嫩平原是世界三大鹽堿地之一,約有鹽堿地373.33萬hm2,這嚴重制約我國東北地區農業系統的正常發展,限制糧食產量。然而,隨著我國人口的持續增長和可耕種土壤的退化,合理開發利用鹽堿地是增加我國糧食產量的重要舉措之一,也是實現農業可持續發展的關鍵因素之一。隨著功能基因組學、分子生物學和基因工程技術日新月異的發展,通過轉基因分子育種改良作物耐鹽堿性,已成為改良和合理開發利用鹽堿地的手段之一。然而,其關鍵在于挖掘功能顯著的耐鹽堿關鍵調控基因。
在生命有機體的生長發育過程中,一系列的生理生化反應都需要在特定ph條件下進行,如細胞質ph維持在7-8。質膜h+-atpase在植物和真菌中通過偶聯atp的水解對質子進行轉運,產生的ph梯度為次級轉運器提供驅動力。植物質膜h+-atpase是一類質子泵,在植物營養吸收、細胞內ph調節、氣孔開閉和細胞生長等生理過程中是不可或缺的。除此之外,在植物應對外界不利因素也具有重要作用,如鹽堿、低溫、干旱、重金屬毒害、光脅迫、化學脅迫和激素脅迫等非生物脅迫。
技術實現要素:
本發明所要解決的技術問題是如何調控植物耐逆性。
為解決上述技術問題,本發明首先提供了與植物耐逆性相關蛋白質gsha12的新用途。
本發明提供了蛋白質gsha12在調控植物耐逆性中的應用。
本發明所涉及的蛋白質gsha12是如下a)或b)或c)或d)的蛋白質:
a)氨基酸序列是序列2所示的蛋白質;
b)在序列2所示的蛋白質的n端和/或c端連接標簽得到的融合蛋白質;
c)將序列2所示的氨基酸序列經過一個或幾個氨基酸殘基的取代和/或缺失和/或添加得到的具有相同功能的蛋白質;
d)與序列2所示的氨基酸序列具有75%或75%以上的同源性且具有相同功能的蛋白質。
其中,序列2由888個氨基酸殘基組成。
為了解決上述技術問題,本發明還提供了與蛋白質gsha12相關的生物材料的新用途。
本發明提供了與蛋白質gsha12相關的生物材料在調控植物耐逆性中的應用。
所述與蛋白質gsha12相關的生物材料為下述a1)至a12)中的任一種:
a1)編碼蛋白質gsha12的核酸分子;
a2)含有a1)所述核酸分子的表達盒;
a3)含有a1)所述核酸分子的重組載體;
a4)含有a2)所述表達盒的重組載體;
a5)含有a1)所述核酸分子的重組微生物;
a6)含有a2)所述表達盒的重組微生物;
a7)含有a3)所述重組載體的重組微生物;
a8)含有a4)所述重組載體的重組微生物;
a9)含有a1)所述核酸分子的轉基因植物細胞系;
a10)含有a2)所述表達盒的轉基因植物細胞系;
a11)含有a3)所述重組載體的轉基因植物細胞系;
a12)含有a4)所述重組載體的轉基因植物細胞系。
上述應用中,a1)所述核酸分子為如下1)或2)或3)所示的基因:
1)其編碼序列是序列1所示的cdna分子或基因組dna分子;
2)與1)限定的核苷酸序列具有75%或75%以上同一性,且編碼蛋白質gsha12的cdna分子或基因組dna分子;
3)在嚴格條件下與1)或2)限定的核苷酸序列雜交,且編碼蛋白質gsha12的cdna分子或基因組dna分子。
其中,所述核酸分子可以是dna,如cdna、基因組dna或重組dna;所述核酸分子也可以是rna,如mrna或hnrna等。
其中,序列1由2667個核苷酸組成,編碼序列2所示的氨基酸序列。
本領域普通技術人員可以很容易地采用已知的方法,例如定向進化和點突變的方法,對本發明的編碼蛋白質gsha12的核苷酸序列進行突變。那些經過人工修飾的,具有編碼蛋白質gsha12的核苷酸序列75%或者更高同一性的核苷酸,只要編碼蛋白質gsha12且具有相同功能,均是衍生于本發明的核苷酸序列并且等同于本發明的序列。
這里使用的術語“同一性”指與天然核酸序列的序列相似性。“同一性”包括與本發明的編碼序列2所示的氨基酸序列組成的蛋白質的核苷酸序列具有75%或更高,或85%或更高,或90%或更高,或95%或更高同一性的核苷酸序列。同一性可以用肉眼或計算機軟件進行評價。使用計算機軟件,兩個或多個序列之間的同一性可以用百分比(%)表示,其可以用來評價相關序列之間的同一性。
上述75%或75%以上同一性,可為80%、85%、90%或95%以上的同一性。
上述應用中,所述嚴格條件是在2×ssc,0.1%sds的溶液中,在68℃下雜交并洗膜2次,每次5min,又于0.5×ssc,0.1%sds的溶液中,在68℃下雜交并洗膜2次,每次15min;或,0.1×sspe(或0.1×ssc)、0.1%sds的溶液中,65℃條件下雜交并洗膜。
上述應用中,a2)所述的含有編碼蛋白質gsha12的核酸分子的表達盒(gsha12基因表達盒)是指能夠在宿主細胞中表達蛋白質gsha12的dna,該dna不但可包括啟動gsha12轉錄的啟動子,還可包括終止gsha12轉錄的終止子。進一步,所述表達盒還可包括增強子序列。可用于本發明的啟動子包括但不限于:組成型啟動子;組織、器官和發育特異的啟動子及誘導型啟動子。合適的轉錄終止子包括但不限于:農桿菌胭脂堿合成酶終止子(nos終止子)、花椰菜花葉病毒camv35s終止子、tml終止子、豌豆rbcse9終止子和胭脂氨酸和章魚氨酸合酶終止子。
可用現有的表達載體構建含有所述gsha12基因表達盒的重組載體。所述植物表達載體包括雙元農桿菌載體和可用于植物微彈轟擊的載體等。如pahc25、pbin438、pcambia1302、pcambia2301、pcambia1301、pcambia1300、pbi121、pcambia1391-xa或pcambia1391-xb(cambia公司)等。所述植物表達載體還可包含外源基因的3′端非翻譯區域,即包含聚腺苷酸信號和任何其它參與mrna加工或基因表達的dna片段。所述聚腺苷酸信號可引導聚腺苷酸加入到mrna前體的3′端,如農桿菌冠癭瘤誘導(ti)質粒基因(如胭脂堿合成酶基因nos)、植物基因(如大豆貯存蛋白基因)3′端轉錄的非翻譯區均具有類似功能。使用本發明的基因構建植物表達載體時,還可使用增強子,包括翻譯增強子或轉錄增強子,這些增強子區域可以是atg起始密碼子或鄰接區域起始密碼子等,但必需與編碼序列的閱讀框相同,以保證整個序列的正確翻譯。所述翻譯控制信號和起始密碼子的來源是廣泛的,可以是天然的,也可以是合成的。翻譯起始區域可以來自轉錄起始區域或結構基因。為了便于對轉基因植物細胞或植物進行鑒定及篩選,可對所用植物表達載體進行加工,如加入可在植物中表達的編碼可產生顏色變化的酶或發光化合物的基因(gus基因、螢光素酶基因等)、抗生素的標記基因(如賦予對卡那霉素和相關抗生素抗性的nptii基因,賦予對除草劑膦絲菌素抗性的bar基因,賦予對抗生素潮霉素抗性的hph基因,和賦予對氨甲喋呤抗性的dhfr基因,賦予對草甘磷抗性的epsps基因)或是抗化學試劑標記基因等(如抗除莠劑基因)、提供代謝甘露糖能力的甘露糖-6-磷酸異構酶基因。從轉基因植物的安全性考慮,可不加任何選擇性標記基因,直接以逆境篩選轉化植株。
上述應用中,所述載體可為質粒、黏粒、噬菌體或病毒載體。
上述應用中,所述微生物可為酵母、細菌、藻或真菌,如農桿菌。
上述應用中,所述轉基因植物細胞系、轉基因植物組織和轉基因植物器官均不包括繁殖材料。
為了解決上述技術問題,本發明還提供了蛋白質gsha12或上述相關生物材料的新用途。
本發明提供了蛋白質gsha12或上述相關生物材料在培育耐逆性提高的轉基因植物中的應用。
本發明還提供了蛋白質gsha12或上述相關生物材料在植物育種中的應用。
上述應用中,所述耐逆性為耐堿性;所述耐堿性具體為耐碳酸鹽脅迫;所述耐碳酸鹽脅迫具體為耐nahco3脅迫。
上述應用中,所述調控為提高。
上述應用中,所述植物為單子葉植物或雙子葉植物,所述雙子葉植物具體可為豆科植物和/或十字花科植物和/或菊科植物;所述豆科植物可為大豆、百脈根、苜蓿或水黃皮;所述十字花科植物可為擬南芥或油菜;所述菊科植物可為向日葵;所述擬南芥可為擬南芥(哥倫比亞生態型col-0)。
為了解決上述技術問題,本發明最后提供了一種培育耐逆性提高的轉基因植物的方法。
本發明提供的培育耐逆性提高的轉基因植物的方法包括提高受體植物中蛋白質gsha12的表達量和/或活性,得到轉基因植物的步驟;所述轉基因植物的耐逆性高于所述受體植物。
上述方法中,所述耐逆性為耐堿性;所述耐堿性具體為耐碳酸鹽脅迫。
上述方法中,所述耐碳酸鹽脅迫為耐nahco3脅迫,具體體現為在nahco3脅迫的條件下:轉基因植物的種子萌發速率高于受體植物和/或轉基因植物的地上部分的鮮重高于受體植物和/或轉基因植物的長勢優于野生型。所述nahco3的濃度具體可為6mmnahco3或8mmnahco3或150mmnahco3。
上述方法中,所述提高受體植物中蛋白質gsha12的表達量和/或活性的方法為在受體植物中過表達蛋白質gsha12。所述過表達的方法為將蛋白質gsha12的編碼基因導入受體植物。
上述方法中,所述蛋白質gsha12的編碼基因的核苷酸序列是序列1所示的dna分子。在本發明的實施例中,所述蛋白質gsha12的編碼基因(即序列表中序列1所示的dna分子)通過重組載體pcambia330035su-gsha12導入所述受體植物中,所述重組載體pcambia330035su-gsha12為在pcambia330035su載體中插入如序列表中序列1所示的gsha12基因。gsha12基因編碼的蛋白的氨基酸序列如序列表中序列2所示。
上述方法中,所述受體植物為單子葉植物或雙子葉植物,所述雙子葉植物具體可為豆科植物和/或十字花科植物和/或菊科植物;所述豆科植物可為大豆、百脈根、苜蓿或水黃皮;所述十字花科植物可為擬南芥或油菜;所述菊科植物可為向日葵;所述擬南芥可為擬南芥(哥倫比亞生態型col-0)。
上述方法中,所述轉基因植物理解為不僅包含將所述gsha12基因轉化受體植物得到的第一代轉基因植物,也包括其子代。對于轉基因植物,可以在該物種中繁殖該基因,也可用常規育種技術將該基因轉移進入相同物種的其它品種,特別包括商業品種中。所述轉基因植物包括種子、愈傷組織、完整植株和細胞。
擴增編碼上述蛋白質gsha12的核酸分子全長或其片段的引物對也屬于本發明的保護范圍。
本發明將gsha12基因超表達于擬南芥中,得到轉gsha12擬南芥。通過實驗證明:在碳酸鹽脅迫下,轉gsha12擬南芥的種子萌發速率和地上部分的鮮重均高于受體植物。說明gsha12基因可增強擬南芥對碳酸鹽脅迫的耐性,gsha12蛋白可以為培育具有碳酸鹽脅迫耐性的轉基因植物的研究奠定基礎。
附圖說明
圖1為gsha12基因在碳酸鹽脅迫下的表達模式分析。
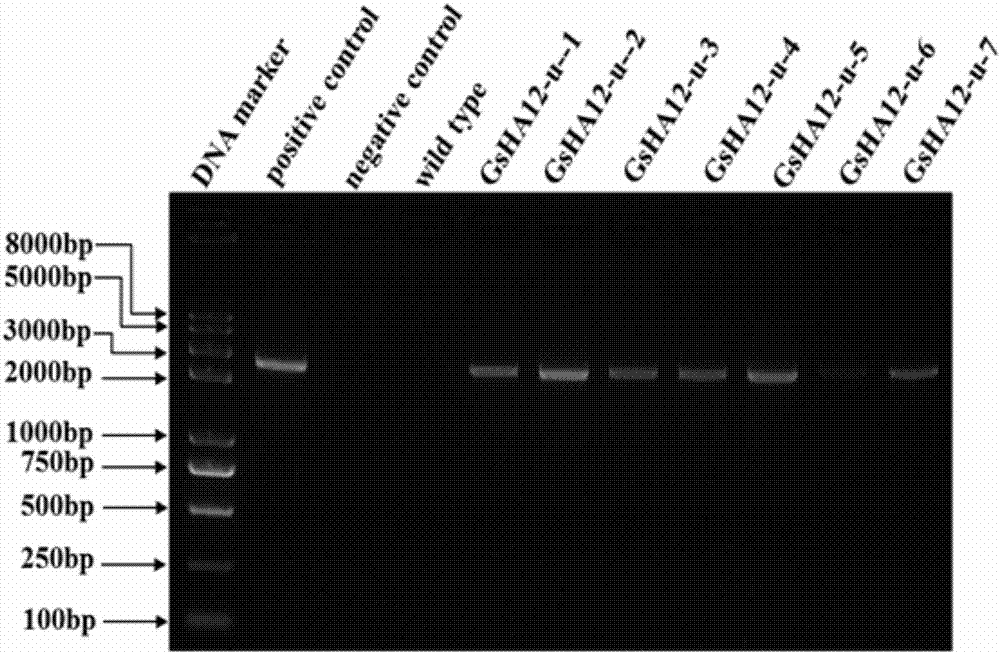
圖2為轉gsha12擬南芥植株的pcr檢測。
圖3為轉gsha12擬南芥植株的rt-pcr檢測。
圖4為轉gsha12擬南芥植株萌發期的耐碳酸鹽脅迫分析。
圖5為轉gsha12擬南芥植株幼苗期的耐碳酸鹽脅迫分析。
圖6為轉gsha12擬南芥植株成苗期的耐碳酸鹽脅迫分析。
具體實施方式
下述實施例中所使用的實驗方法如無特殊說明,均為常規方法。
下述實施例中所用的材料、試劑等,如無特殊說明,均可從商業途徑得到。
下述實施例中的定量試驗,均設置三次重復實驗,結果取平均值。
下述實施例中的野生型大豆g07256記載于如下文獻:mingzhesun,xiaolisun,yangzhao,huacai,chaoyuezhao,weiji,huiziduanmu,yangyu,yanmingzhu.ectopicexpressionofgsppck3andscmrpinmedicagosativaenhancesplantalkalinestresstoleranceandmethioninecontent.plosone2014,9(2):e89578,公眾可從申請人(黑龍江八一農墾大學)處獲得,該生物材料只為重復本發明的相關實驗所用,不可作為其它用途使用。
下述實施例中的pcambia330035su記載于如下文獻:xiaolisun,weiji,xiaodongding,xibai,huacai,shanshanyang,xueqian,mingzhesun,yanmingzhu.gsvamp72,anovelglycinesojar-snareprotein,isinvolvedinregulatingplantsalttoleranceandabasensitivity.plantcelltissorgancult2013,113:199–215,公眾可從申請人(黑龍江八一農墾大學)處獲得,該生物材料只為重復本發明的相關實驗所用,不可作為其它用途使用。
下述實施例中的根癌農桿菌gv3101記載于如下文獻:leecw,etal.agrobacteriumtumefacienspromotestumorinductionbymodulatingpathogendefenseinarabidopsisthaliana,plantcell,2009,21(9),2948-62,公眾可從申請人(黑龍江八一農墾大學)處獲得,該生物材料只為重復本發明的相關實驗所用,不可作為其它用途使用。
下述實施例中的野生型擬南芥(哥倫比亞生態型col-0)記載于如下文獻:luox,sunx,liub,etal.ectopicexpressionofawrkyhomologfromglycinesojaaltersfloweringtimeinarabidopsis[j].plosone,2013,8(8):e73295,公眾可從申請人(黑龍江八一農墾大學)處獲得,該生物材料只為重復本發明的相關實驗所用,不可作為其它用途使用。
實施例1、野生大豆gsha12全長基因的克隆
1、總rna的提取
選取飽滿的野生大豆g07256種子,用濃h2so4處理10min,無菌水沖洗3-4次,25℃暗培養2-3d催芽。待芽長到1-2cm時,將其轉移至1/4hogland營養液中,置于人工氣候箱中培養。取3周齡野生大豆g07256幼苗的根,采用rnapreppure試劑盒(tiangen)提取總rna。
2、cdna的獲得
以oligodt為引物進行cdna第一條鏈的合成,方法參見invitrogen公司superscripttmiiireversetranscriptase說明書,得到野生大豆總cdna。
3、pcr擴增
以野生大豆總cdna為模板,采用引物gsha12-s和gsha12-as進行pcr擴增,得到pcr擴增產物。引物序列如下:
gsha12-s:5'-atgtggaatcctctttcatgggtca-3';
gsha12-as:5'-ctagacagtgtatgcttgctgtattgtgtc-3';
4、gsha12基因的克隆載體構建及測序
將pcr擴增產物與peasy-t載體連接,構建peasy-gsha12克隆載體。并對其進行測序。
測序結果表明:pcr擴增得到的大小為2667bp的dna片段,其核苷酸序列如序列表中序列1所示,將序列1所示的基因命名為gsha12基因,自5’端第1-2667位為orf,gsha12基因編碼序列表中序列2所示的蛋白質,將序列2所示的氨基酸序列命名為gsha12蛋白。
實施例2、gsha12基因在碳酸鹽脅迫條件下的表達模式分析
1、對3周齡的野生大豆g07256幼苗進行50mmnahco3碳酸鹽脅迫處理,分別在處理0h、1h、3h、6h、12h、24h時取根尖組織。
2、采用rna提取試劑盒rneasyplantminikit(qiagen,hilden,germany)提取總rna,采用反轉錄試劑盒superscripttmiiireversetranscriptasekit(invitrogen,carlsbad,ca,usa)反轉錄獲得cdna。
3、采用sybr定量試劑盒sybrpremixextaqtmiimix(takara,shiga,japan)于熒光定量pcr儀abi7500(appliedbiosystems,usa)上進行real-timepcr檢測基因表達量。定量分析采用比較ct法(△△ct),以gapdh基因為內參,未經處理的樣品作為對照。經gapdh基因均一化處理后,通過2-△△ct法計算gsha12基因表達量變化差異,以處理樣本相對于未處理樣本的倍數表示。gsha12基因特異引物為5’-aggcaagggctggtattcaagag-3’和5’-gagaccgtaatcctcgctctgca-3’,gapdh基因特異引物為5’-gactggtatggcattccgtgt-3’和5’-gccctctgattcctccttga-3’。
定量real-timepcr結果顯示碳酸鹽脅迫處理后,gsha12基因表達量呈上升趨勢,并在碳酸鹽脅迫處理12h后達到頂峰(圖1),表明gsha12基因的表達受碳酸鹽脅迫誘導。
實施例3、轉gsha12擬南芥的獲得及耐碳酸鹽分析
一、轉gsha12擬南芥的獲得
1、以peasy-gsha12克隆載體質粒為模板,采用基因特異引物gsha12-u-f和gsha12-u-r進行pcr擴增,得到gsha12基因全長cds區。引物序列如下(下劃線代表載體構建時所需的接頭序列,其中u為user酶切位點):
gsha12-u-f:5’-ggcttaauatgtggaatcctctttcat-3’;
gsha12-u-r:5’-ggtttaauctagacagtgtatgcttgctg-3’。
2、用限制性內切酶paci和nt.bbvci對pcambia330035su載體進行雙酶切,得到載體酶切產物。將獲得的載體酶切產物、user酶(neb,m5505s)和步驟1獲得的gsha12基因在37℃下孵育20min,利用user酶對gsha12基因片段的尿嘧啶處進行切割,形成可與pcambia330035su載體互補的粘性末端,接著25℃下孵育20min,得到重組表達載體pcambia330035su-gsha12,并送交測序。
測序結果表明:重組表達載體pcambia330035su-gsha12為在pcambia330035su載體中插入如序列表中序列1所示的gsha12基因,其保持pcambia330035su載體的其他序列不變后得到的載體。重組表達載體pcambia330035su-gsha12表達的gsha12蛋白。
3、采用凍融法,將pcambia330035su-gsha12載體轉化至根癌農桿菌gv3101,獲得重組農桿菌,并經pcr鑒定得到陽性轉化子(含有序列表中序列1所示的gsha12轉化子),用于侵染擬南芥植株。
4、轉gsha12擬南芥的獲得及鑒定
將上述重組農桿菌通過floral-dip法侵染野生型擬南芥(哥倫比亞生態型)。并將t0代種子表面消毒后,播種于含25mg/l固殺草的1/2ms培養基上進行篩選。將t1代抗性苗移栽至營養缽中培養,提取基因組dna,進行pcr鑒定和rt-pcr鑒定。
(1)pcr鑒定
采用easypureplantgenomicdnakit基因組提取試劑盒(全式金,ee111-01),提取野生型擬南芥植株和固殺草抗性植株的基因組dna,以獲得的基因組dna為模板,并采用5'-atgtggaatcctctttcatgggtca-3'和5'-ctagacagtgtatgcttgctgtattgtgtc-3'引物進行pcr鑒定。結果如圖2所示。從圖中可以看出:抗性植株gsha12-u-1、gsha12-u-2、gsha12-u-3、gsha12-u-4、gsha12-u-5、gsha12-u-6和gsha12-u-7均擴增得到目的條帶,而野生型擬南芥植株沒有擴增得到的目的條帶。
(2)rt-pcr鑒定
對pcr鑒定陽性的植株,提取總rna,利用實施例1中的real-timepcr引物(5’-aggcaagggctggtattcaagag-3’和5’-gagaccgtaatcctcgctctgca-3’)進行半定量rt-pcr,以擬南芥actin2基因為內參,檢測gsha12基因在轉基因植株中的表達量。actin2基因特異引物序列如下:actin2-rt-f:5’-ttacccgatgggcaagtc-3’;actin2-rt-r:5’-gctcatacggtcagcgatac-3’。
結果如圖3所示。從圖中可以看出:野生型擬南芥植株的rt-pcr無擴增產物,而轉gsha12擬南芥均可以擴增出目的條帶,表明外源基因gsha12不但已順利整合到擬南芥的基因組上,而且能夠在轉基因擬南芥中正常轉錄表達。
將rt-pcr陽性的t1代轉gsha12擬南芥單株收種子,并分別播種于含25mg/l固殺草的1/2ms培養基上進行篩選,觀察t2代分離情況。如此重復,直至得到t3代轉gsha12擬南芥純合體株系。選取t3代轉gsha12擬南芥純合體株系(#1)和(#2)用于下述耐碳酸鹽分析。
二、轉gsha12擬南芥的耐碳酸鹽分析
1、轉gsha12擬南芥萌發期的耐碳酸鹽分析
選取飽滿的野生型擬南芥、t3代轉gsha12擬南芥純合體株系(#1)和(#2)種子,用5%naclo處理6-8min,滅菌ddh2o沖洗5-7次,4℃春化3d,一部分播種于正常的1/2ms培養基,一部分播種于含6mmnahco3的1/2ms培養基,22℃培養3d,統計種子萌發率。實驗重復三次,每種處理各株系采用30株植株。
結果如圖4所示。從圖中可以看出:碳酸鹽脅迫嚴重抑制了野生型與轉gsha12擬南芥種子的萌發,但轉gsha12擬南芥純合體株系(#1)和(#2)的種子萌發率高于野生型。
2、轉gsha12擬南芥幼苗期的耐碳酸鹽分析
選取飽滿的野生型擬南芥、t3代轉gsha12擬南芥純合體株系(#1)和(#2)種子,春化后播種于正常的1/2ms培養基,22℃培養11d。待擬南芥長至六葉期,將幼苗移至正常、或含8mmnahco3的1/2ms培養基豎直培養7d。實驗重復三次,每種處理各株系采用15株植株。
結果如圖5所示。從圖中可以看出:碳酸鹽脅迫抑制了野生型和轉gsha12擬南芥根的伸長和地上部分的生長,并且t3代轉gsha12擬南芥純合體株系(#1)和(#2)地上部分的鮮重也顯著高于野生型。
3、轉gsha12擬南芥成苗期的耐碳酸鹽分析
選取飽滿的野生型擬南芥、t3代轉gsha12擬南芥純合體株系(#1)和(#2)種子,春化后播種于營養缽中(營養土:君子蘭土:蛭石1:1:1),置于人工氣候培養箱中培養。選取長勢一致的4周齡擬南芥植株,每3d澆灌1次150mmnahco3(ph9.0)溶液進行碳酸鹽脅迫處理,觀察脅迫處理后植株長勢。實驗重復三次,每種處理各株系采用30株植株。
結果如圖6所示。從圖中可以看出:碳酸鹽脅迫處理后野生型和轉gsha12擬南芥都逐漸失綠發黃,變紫甚至死亡,但是t3代轉gsha12擬南芥純合體株系(#1)和(#2)的長勢明顯優于野生型。
上述結果表明gsha12基因超量表達顯著提高了植物的耐逆性,尤其是碳酸鹽脅迫耐性。
序列表
<110>黑龍江八一農墾大學
<120>gsha12蛋白及其編碼基因在調控植物耐逆性中的應用
<160>2
<210>1
<211>2667bp
<212>dna
<213>野生大豆(glycinesoja)
<400>1
atgtggaatcctctttcatgggtcatggaagctgcagcaatcatggccattgctttggcc60
aatggaggaggaaaacctcctgattggcaagactttgttgggattatcacactccttatt120
atcaattcaacaataagtttcattgaggagaacaatgctggtaatgctgcggcagctctg180
atggctcgtttagcacctaaagctaagttccttcgagatgggaaatggattgaggaggat240
gctagcattcttgttcctggtgatataattagtgttaagctaggggatattatccctgcg300
gatgctcgtctacttgaaggtgatccactgaagattgatcagtctgcacttacaggcgag360
tctcttcctgtcacaaaaggccctggtgatagtgtttattcaggctccacatgcaagcag420
ggagagatcaatgcagttgttattgccacaggagttcataccttctttggcaaagctgct480
catcttgtggactccacaaatcaagttggtcatttccagaaggtcctgactgcaattggg540
aacttctgcatatgttccattgctgtgggaatgatagtagagataattgtcatgtaccca600
attcaacaccgggaatatcgtcctgggattgacaatctgcttgtgcttcttattggagga660
attcctattgccatgcctactgttttgtcagtgacaatggcaattggatcccatcgctta720
gctcagcagggtgctattactaaaagaatgacagcaatagaagagatggcaggaatggat780
gtattatgtagtgacaaaactggaactttgactttgaataaactgacagttgacaagaat840
cttattgagatttttgctaaaggagttgacgtagatactgttgttctcatggccgctcgg900
gctgcacgattggaaaaccaagatgctatagatgccgctattgtagggatgttgggtgat960
ccaaaagaggcaagggctggtattcaagaggttcacttcctacccttcaatccaactgac1020
aagcgaactgcaatcacttatatagacggtgaaagtaaaatgcatcgtgtcagcaaagga1080
gcaccagagcagattttgaatcttgcacgcaataaatcagagatagaacgcagagttcat1140
tctgtcattgataagtttgcagagcgaggattacggtctcttgcagtagcttaccaggaa1200
gttcctgatggaaagaaagaaagccaaggagggccttggcaatttattggactgttgcct1260
ttatttgacccacctagacatgatagtgctgagacaatacgaagggcattaaatcttgga1320
gtaaatgttaaaatgataacaggtgatcaactagcaataggaaaagaaacaggacgccgt1380
ctggggatgggaaccaacatgtacccttcatcggctttattgggccaaaacaaggatgaa1440
gcaattgctaccttgccagttgatgagttgattgaaaaagcagatggatttgctggtgtt1500
tttcctgaacacaaatatgagatcgtgaaacgtttacaagctaggaaacacatatgtgga1560
atgactggtgatggggttaacgatgctcctgctcttaaaaaggcagatattggaatagct1620
gtcgccgatgctactgatgcagctcgtagtgcttctgatattgttctgactgaacctggt1680
ctcagtgttatcatcagtgctgtactgaccagtcgagcaatattccaaaggatgaagaat1740
tacacaatctatgcagtttccatcacaatccgtattgtgcttggtttcatgttactggcc1800
ctcatatggcattttgattttccaccattcatggtgctgattattgctattcttaatgac1860
ggtaccattatgacgatatcaaaggacagggtgaaaccatctccatatccagatagctgg1920
aagttggccgagatctttaccactggaataattcttggtggttatttggctatgatgaca1980
gttattttcttttgggcagcatataaaacagattttttccctcaaacatttggagtctca2040
agtcttcagaaaaaggatagggatgactttagaaagcttgcctcagcaatatacctacaa2100
gttagcacaattagtcaggccctcatattcattacacgggctcggagttggtcttatgtt2160
gaacgtccgggtttgttacttgttgcagcttttgttatcgcccagctgatagctacctta2220
attgcagtttatgcaaattggagtttcgctgctattgaagggattggatggggttgggct2280
ggtgttgtttggctttacaacctcatcttttatatcccacttgactttatcaagttcata2340
attcgatatgccttgagtggaagggcttgggatcttgttattgaacaaaggattgctttt2400
acaaggaaaaaagattttggaaaggaagaacgtgaacttaaatgggcacatgcacagagg2460
acgcttcacggccttcacccaccagagactaagatgttcaatgaacgtacaagttacaca2520
gaacttaatcagatggctgaagaggctagaagacgagcagaaattgcaaggctgagagaa2580
ctgcatacacttaagggtcgtgttgagtctgtggttagactgaagggtcttaacattgac2640
acaatacagcaagcatacactgtctag2667
<210>2
<211>888
<212>prt
<213>野生大豆(glycinesoja)
<400>2
mettrpasnproleusertrpvalmetglualaalaalailemetala
151015
ilealaleualaasnglyglyglylysproproasptrpglnaspphe
202530
valglyileilethrleuleuileileasnserthrileserpheile
354045
glugluasnasnalaglyasnalaalaalaalaleumetalaargleu
505560
alaprolysalalyspheleuargaspglylystrpileglugluasp
65707580
alaserileleuvalproglyaspileileservallysleuglyasp
859095
ileileproalaaspalaargleuleugluglyaspproleulysile
100105110
aspglnseralaleuthrglygluserleuprovalthrlysglypro
115120125
glyaspservaltyrserglyserthrcyslysglnglygluileasn
130135140
alavalvalilealathrglyvalhisthrphepheglylysalaala
145150155160
hisleuvalaspserthrasnglnvalglyhispheglnlysvalleu
165170175
thralaileglyasnphecysilecysserilealavalglymetile
180185190
valgluileilevalmettyrproileglnhisargglutyrargpro
195200205
glyileaspasnleuleuvalleuleuileglyglyileproileala
210215220
metprothrvalleuservalthrmetalaileglyserhisargleu
225230235240
alaglnglnglyalailethrlysargmetthralailegluglumet
245250255
alaglymetaspvalleucysserasplysthrglythrleuthrleu
260265270
asnlysleuthrvalasplysasnleuilegluilephealalysgly
275280285
valaspvalaspthrvalvalleumetalaalaargalaalaargleu
290295300
gluasnglnaspalaileaspalaalailevalglymetleuglyasp
305310315320
prolysglualaargalaglyileglngluvalhispheleuprophe
325330335
asnprothrasplysargthralailethrtyrileaspglygluser
340345350
lysmethisargvalserlysglyalaprogluglnileleuasnleu
355360365
alaargasnlyssergluilegluargargvalhisservalileasp
370375380
lysphealagluargglyleuargserleualavalalatyrglnglu
385390395400
valproaspglylyslysgluserglnglyglyprotrpglnpheile
405410415
glyleuleuproleupheaspproproarghisaspseralagluthr
420425430
ileargargalaleuasnleuglyvalasnvallysmetilethrgly
435440445
aspglnleualaileglylysgluthrglyargargleuglymetgly
450455460
thrasnmettyrproserseralaleuleuglyglnasnlysaspglu
465470475480
alailealathrleuprovalaspgluleuileglulysalaaspgly
485490495
phealaglyvalpheprogluhislystyrgluilevallysargleu
500505510
glnalaarglyshisilecysglymetthrglyaspglyvalasnasp
515520525
alaproalaleulyslysalaaspileglyilealavalalaaspala
530535540
thraspalaalaargseralaseraspilevalleuthrgluprogly
545550555560
leuservalileileseralavalleuthrserargalailephegln
565570575
argmetlysasntyrthriletyralavalserilethrileargile
580585590
valleuglyphemetleuleualaleuiletrphispheaspphepro
595600605
prophemetvalleuileilealaileleuasnaspglythrilemet
610615620
thrileserlysaspargvallysproserprotyrproaspsertrp
625630635640
lysleualagluilephethrthrglyileileleuglyglytyrleu
645650655
alametmetthrvalilephephetrpalaalatyrlysthraspphe
660665670
pheproglnthrpheglyvalserserleuglnlyslysaspargasp
675680685
aspphearglysleualaseralailetyrleuglnvalserthrile
690695700
serglnalaleuilepheilethrargalaargsertrpsertyrval
705710715720
gluargproglyleuleuleuvalalaalaphevalilealaglnleu
725730735
ilealathrleuilealavaltyralaasntrpserphealaalaile
740745750
gluglyileglytrpglytrpalaglyvalvaltrpleutyrasnleu
755760765
ilephetyrileproleuasppheilelyspheileileargtyrala
770775780
leuserglyargalatrpaspleuvalilegluglnargilealaphe
785790795800
thrarglyslysasppheglylysglugluarggluleulystrpala
805810815
hisalaglnargthrleuhisglyleuhisproprogluthrlysmet
820825830
pheasngluargthrsertyrthrgluleuasnglnmetalagluglu
835840845
alaargargargalagluilealaargleuarggluleuhisthrleu
850855860
lysglyargvalgluservalvalargleulysglyleuasnileasp
865870875880
thrileglnglnalatyrthrval
885
- 還沒有人留言評論。精彩留言會獲得點贊!
- 蝎神經毒素AaIT及其編碼基因與應用的制造方法與工藝
- 腈水解酶突變體及其編碼基因和應用的制造方法與工藝
- 植物抗旱相關蛋白AsTMK1及其編碼基因和應用的制造方法與工藝
- 植物抗旱相關蛋白EtSnRK2.2及其編碼基因和應用的制造方法與工藝
- OsSAPK7蛋白及其編碼基因在提高水稻白葉枯病抗性中的應用的制造方法與工藝
- 一種抗除草劑抗蟲融合基因、編碼蛋白及應用的制造方法與工藝
- 毛果楊PtrZFP103基因及其編碼蛋白和應用的制造方法與工藝
- 大豆bHLH轉錄因子基因GmFER及其編碼蛋白和應用的制造方法與工藝
- 一種花狹口蛙的基因及其編碼的多肽和該多肽的用途的制造方法與工藝
- 一種澤陸蛙基因及其編碼的多肽和該多肽的用途的制造方法與工藝
- 降低種子重量的白樺AP2基因及其編碼蛋白的制造方法與工藝
- 可編碼ArtRD蛋白基因及其蛋白和應用的制造方法與工藝
- 蝎神經毒素AaIT及其編碼基因與應用的制造方法與工藝
- 腈水解酶突變體及其編碼基因和應用的制造方法與工藝
- 植物抗旱相關蛋白AsTMK1及其編碼基因和應用的制造方法與工藝
- 植物抗旱相關蛋白EtSnRK2.2及其編碼基因和應用的制造方法與工藝
- OsSAPK7蛋白及其編碼基因在提高水稻白葉枯病抗性中的應用的制造方法與工藝
- 一種抗除草劑抗蟲融合基因、編碼蛋白及應用的制造方法與工藝
- 毛果楊PtrZFP103基因及其編碼蛋白和應用的制造方法與工藝
- 大豆bHLH轉錄因子基因GmFER及其編碼蛋白和應用的制造方法與工藝
- 腈水解酶突變體及其編碼基因和應用的制造方法與工藝
- 植物抗旱相關蛋白AsTMK1及其編碼基因和應用的制造方法與工藝
- 植物抗旱相關蛋白EtSnRK2.2及其編碼基因和應用的制造方法與工藝
- OsSAPK7蛋白及其編碼基因在提高水稻白葉枯病抗性中的應用的制造方法與工藝
- 一種抗除草劑抗蟲融合基因、編碼蛋白及應用的制造方法與工藝
- 毛果楊PtrZFP103基因及其編碼蛋白和應用的制造方法與工藝
- 大豆bHLH轉錄因子基因GmFER及其編碼蛋白和應用的制造方法與工藝
- 一種花狹口蛙的基因及其編碼的多肽和該多肽的用途的制造方法與工藝
- 一種澤陸蛙基因及其編碼的多肽和該多肽的用途的制造方法與工藝
- 一種水稻MIT1基因、其編碼蛋白及應用的制造方法與工藝
- 植物抗旱相關蛋白EtSnRK2.2及其編碼基因和應用的制造方法與工藝
- OsSAPK7蛋白及其編碼基因在提高水稻白葉枯病抗性中的應用的制造方法與工藝
- 一種抗除草劑抗蟲融合基因、編碼蛋白及應用的制造方法與工藝
- 毛果楊PtrZFP103基因及其編碼蛋白和應用的制造方法與工藝
- 大豆bHLH轉錄因子基因GmFER及其編碼蛋白和應用的制造方法與工藝
- 一種水稻MIT1基因、其編碼蛋白及應用的制造方法與工藝
- 蛋白StTrxF、編碼基因及其在提高植物淀粉含量中的應用的制造方法與工藝
- 蛋白質及其編碼基因在調控植物抗黃萎病性中的應用的制造方法與工藝
- 甘薯耐逆相關蛋白IbFBA2、編碼基因、重組載體與應用的制造方法與工藝
- 植物抗旱相關蛋白ZmNAC111及其編碼基因與應用的制造方法與工藝
- 植物抗旱相關蛋白EtSnRK2.2及其編碼基因和應用的制造方法與工藝
- OsSAPK7蛋白及其編碼基因在提高水稻白葉枯病抗性中的應用的制造方法與工藝
- 一種抗除草劑抗蟲融合基因、編碼蛋白及應用的制造方法與工藝
- 毛果楊PtrZFP103基因及其編碼蛋白和應用的制造方法與工藝
- 大豆bHLH轉錄因子基因GmFER及其編碼蛋白和應用的制造方法與工藝
- 一種水稻MIT1基因、其編碼蛋白及應用的制造方法與工藝
- 蛋白StTrxF、編碼基因及其在提高植物淀粉含量中的應用的制造方法與工藝
- 植物抗旱相關蛋白ZmNAC111及其編碼基因與應用的制造方法與工藝
- 核盤菌凝集素SSL?6蛋白及其編碼基因與應用的制造方法與工藝
- 植物抗病毒相關蛋白GmSN1及其編碼基因和應用的制造方法與工藝
- 植物抗旱相關蛋白EtSnRK2.2及其編碼基因和應用的制造方法與工藝
- OsSAPK7蛋白及其編碼基因在提高水稻白葉枯病抗性中的應用的制造方法與工藝
- 一種抗除草劑抗蟲融合基因、編碼蛋白及應用的制造方法與工藝
- 毛果楊PtrZFP103基因及其編碼蛋白和應用的制造方法與工藝
- 大豆bHLH轉錄因子基因GmFER及其編碼蛋白和應用的制造方法與工藝
- 一種水稻MIT1基因、其編碼蛋白及應用的制造方法與工藝
- 蛋白StTrxF、編碼基因及其在提高植物淀粉含量中的應用的制造方法與工藝
- 蛋白質及其編碼基因在調控植物抗黃萎病性中的應用的制造方法與工藝
- 植物抗旱相關蛋白ZmNAC111及其編碼基因與應用的制造方法與工藝
- 核盤菌凝集素SSL?6蛋白及其編碼基因與應用的制造方法與工藝