一種水稻MIT1基因、其編碼蛋白及應用的制作方法
本發明屬于基因工程領域,具體地說,涉及一種水稻MIT1基因、其編碼的蛋白及其在抑制水稻分枝中的應用。
背景技術:
水稻(Oryza sativa)為禾本科單子葉植物,是世界上重要的糧食作物之一。目前,我國水稻種植面積約4.5億畝,居世界第二位,總產量2.04億噸,居世界第一位,單產420公斤,是世界平均水平的1.6倍。然而,隨著人口的持續增長和耕地面積的不斷減少,對糧食生產帶來了更大的挑戰。因此,通過品種的遺傳改良,挖掘作物的單產潛力是解決糧食安全問題的有效途徑,也是保障世界糧食產量穩步增長的根本出路所在。
分蘗是決定水稻產量的重要農藝性狀之一,是單子葉植物生長發育過程中形成的一種特殊的分枝性狀,存在著一定的規律。水稻分蘗發生的早晚或多少,與品種特性有關,分蘗增長速度在品種間也存在差異,即遺傳因素是影響分蘗動態變化的主要原因之一。水稻分蘗分為有效分蘗和無效分蘗,有效分蘗的比例高,即分蘗的成穗率高是群體質量的重要標志。如果能控制最高分蘗數,使后期無效分蘗減少,分蘗成穗率便可大幅度提高,而有望提高產量。分蘗角度為抽穗前主莖與分蘗之間的夾角。分蘗角度過小,常規種植會使植株間的空隙較大,葉面積指數較小,浪費光能;分蘗角度過大,易使個體間遮擋,光能利用率降低。因此,分蘗角度對于水稻株型的構成具有重要意義,與植物的形態,群體密度,光能利用率和風速等水稻生理生態特性有重大的關系。從育種角度來看,豐產性好的水稻品種表現為分蘗力強,成穗率高,穗粒數多,結實率高,這也是實現超級稻高產和優質的保證,即在超級稻生物量較高的基礎上提高收獲指數。選育分蘗變化平穩,有效分蘗期長,成穗率高的品種,產生足夠的群體數量,保證良好的群體結構,更好的協調群體與個體間的矛盾,從而進一步提高產量。
水稻矮源是帶有矮稈或半矮稈基因的種質資源,大量實驗證明矮化能明顯提高水稻的產量。目前生產上應用的矮稈基因主要是sd1。系統分析我國育成的秈稻品種表明,利用的矮稈資源主要有六個:矮腳南特、矮仔占、低腳烏尖、花龍水田谷、矮腳水谷田和中山無名種等。其中矮仔占、矮腳南特、低腳烏尖、花龍水田谷及其衍生品種都是由一對隱性半矮桿基因sd1控制,同時還存在著不同程度的微效多基因的影響。
但是,矮稈基因利用單一和遺傳背景狹隘可能造成遺傳上的脆弱性,近幾年來國內外新育成的許多矮稈品種產量潛力停滯不前也可能與此有關。而已鑒定出的許多半矮稈和矮稈基因不少都對水稻的農藝性狀表現負面的影響,限制了它們在雜交育種中的應用。囚此,發掘和鑒定控制水稻株高的基因,開展株高基因定位、克隆及作用機理等方面的研究,實現對水稻株高的定向改良,具有十分重要的理論意義和應用價值。同時,水稻是研究禾本科植物的模式作物,水稻基因組的測序已完成,這為水稻基因的克隆及其功能提供了極為有利的條件。
研究人員從環境因素、生理生化及基因組學等多方面對植物矮化多分枝機制進行了研究,并取得了很大的進展。從不同的多分枝突變體中發現了一種新的調控植物分枝的植物激素—獨腳金內酯(Strigolactones),它在植物根部產生并向上運輸,從而抑制腋芽的發育和植物的分枝,對于控制植物株型具有重要的意義。獨腳金內酯途徑基因的突變通常會導致植物分蘗或分枝數目的增加,同時伴有植株高度的降低。研究人員從矮化多分枝/分蘗突變體中克隆了多個突變基因,如從腋生分枝增多的擬南芥突變體中克隆了MAX1至MAX4基因,從多分枝的豌豆ramous(rms)突變體中克隆了RMS1至RMS5基因,從矮化多分蘗的水稻突變體dwarf/high tillering dwarf(htd)中克隆了D3、D10、D14/D88/HTD2、D17/HTD1、D27、D53、OsTB1/FC1等基因。從頂端優勢削弱的矮牽牛dad(decreased apical dominance)突變體中克隆的DAD1至DAD3也位于獨腳金內酯途徑中。
獨腳金內酯的生物合成起源于類胡蘿卜素。在植物類胡蘿卜素合成過程中,八氫番茄紅素合成酶(PSY)的產物—15-順式-八氫番茄紅素,通過脫氫反應和異構反應合成下游反應需要的全反式-番茄紅素,被進一步轉化合成胡蘿卜素和葉黃素,作為某些激素的底物,如ABA和SL。在此途徑中,需要兩種異構酶的參與,分別為15-順式-ζ-胡蘿卜素異構酶(Z-ISO)和胡蘿卜素異構酶(CRTISO)。Z-ISO在此途徑中催化9,15,9'-3-順式-ζ-胡蘿卜素發生異構反應,形成9,9'-2-順式-ζ-胡蘿卜素,作為后續反應的的底物。在本發明中,將Z-ISO命名為MIT1。該酶起源于NnrU基因,它編碼一個跨膜蛋白—亞硝酸鹽和一氧化氮還原酶U(nitrite and nitric oxide reductase U,NnrU)。
技術實現要素:
本發明目的是提供水稻MIT1基因及其應用。
本發明首先提供了水稻MIT1蛋白,其具有:
1)如SEQ ID No.1所示的氨基酸序列;或
2)SEQ ID No.1所示的氨基酸序列經取代、缺失和/或增加一個或多個氨基酸且具有同等活性的由1)衍生的蛋白質。
本發明提供了編碼水稻MIT1蛋白的基因,其具有:
1)SEQ ID No.2所示的核苷酸序列;或
2)SEQ ID No.2所示核苷酸序列經取代、缺失和/或增加一個或幾個核苷酸;或
3)在嚴格條件下與1)限定的DNA序列雜交的核苷酸序列。
本發明提供了含有編碼水稻MIT1蛋白的基因的生物材料,所述生物材料為載體、宿主細胞、轉化植物細胞。
本發明提供了水稻MIT1蛋白或其編碼基因在制備轉基因植物中的應用。
本發明提供了水稻MIT1蛋白或其編碼基因在水稻種質資源改良中的應用。
本發明提供了水稻MIT1蛋白或其編碼基因在提高水稻產量中的應用。
本發明還提供一種水稻MIT1基因突變基因,其為水稻MIT1基因第二內含子3’末端的2個堿基AG和第三外顯子5’端的8個堿基GTCTACTT缺失,所述水稻MIT1基因含有如SEQ ID No.2所示的核苷酸序列。
進一步地,所述生物材料為載體、宿主細胞、轉化植物細胞。
本發明提供了上述突變基因或含有該突變基因的生物材料在農作物改良育種、制種中的應用。
所述的農作物為水稻、玉米、小麥或棉花。
本發明提供了上述突變基因或含有該突變基因的生物材料在提高水稻產量中的應用。
本發明的優點在于:
(1)本發明提供了水稻MIT1基因(核苷酸序列如SEQ ID No.2所示)及其編碼的蛋白(氨基酸序列如SEQ ID No.1所示)。將野生型MIT1基因組DNA通過農桿菌介導轉化mit1突變體的愈傷組織,發現互補轉基因植株的表型完全恢復正常,分蘗數和株高都和野生型一致,因而MIT1基因具有抑制水稻分蘗的功能,有望對水稻株型的形成進行調控進而對株型定向設計,以提高水稻生產力,在水稻種質資源改良中篩選效果明顯,經濟價值巨大。
(2)因輻射誘變造成與野生型相比有10bp缺失,采用實驗室常用的瓊脂糖電泳就能開展分子檢測,即能夠實現鑒別,不需要特別的檢測技術和方法。
附圖說明
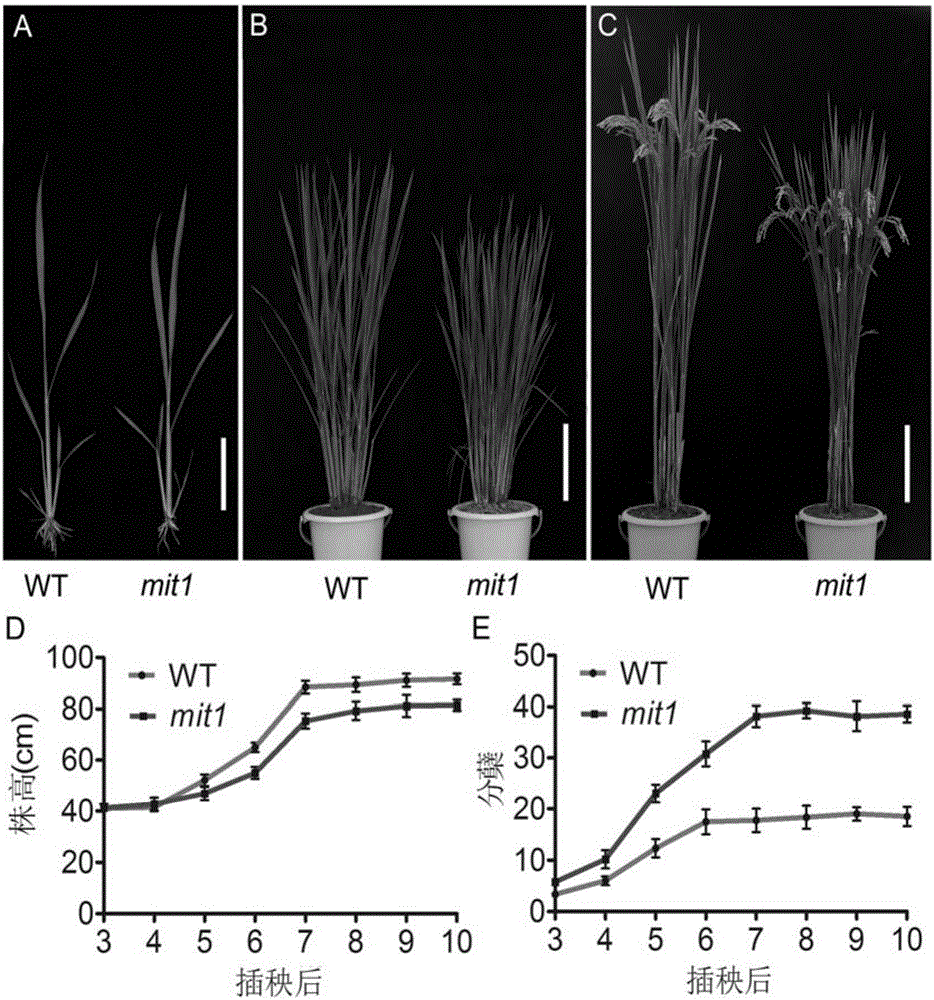
圖1為野生型春江06與MIT1突變體mit1的表型。其中,A,B,C圖分別為野生型和mit1的幼苗期、分蘗期和成熟期的表型;D,E圖分別為插秧后野生型和mit1的株高和分蘗的動態變化。A圖比例尺為5cm,B,C圖比例尺為20cm。
圖2為野生型和mit1的產量相關性狀。A圖為各節間長度比較。比例尺為5cm。B圖為穗形。比例尺為5cm。C圖為粒形。D圖為成熟期株高。E圖為有效分蘗數。F圖為每穗粒數。G圖為百粒重。H圖為單株產量。
圖3為MIT1基因的圖位克隆和功能互補驗證。A圖為MIT1的圖位克隆。B圖為MIT1基因結構及在突變體mit1中的突變位點。C圖為mit1突變體的互補驗證。B260-1和B260-2為兩個轉基因回補株系。比例尺為15cm。
圖4為MIT1的同源蛋白序列比對和進化分析。A圖為水稻、玉米、擬南芥中的MIT1,ZmZ-ISO、AtZ-ISO的蛋白序列比對。B圖為MIT1的進化分析。
圖5為MIT1組織表達模式。
圖6為水稻組織GUS染色結果。圖中,a莖,b莖節,c根,d穗,e葉片,f葉鞘,g側芽,h葉鞘橫切面,i穎殼橫切面,j根橫切面。a,f,g圖的比例尺為200μm,b,c,d,e圖的比例尺為100μm,h,I,j圖的比例尺為50μm。
圖7為MIT1蛋白的亞細胞定位。
具體實施方式
以下實施例用于說明本發明,但不用來限制本發明的范圍。在不背離本發明精神和實質的情況下,對本發明方法、步驟或條件所作的修改或替換,均屬于本發明的范圍。
若未特別指明,實施例中所用的技術手段為本領域技術人員所熟知的常規手段;若未特別指明,實施例中所用試劑均為市售。
實施例1突變體的獲得與表型分析
突變體mit1是經60Co-γ輻射誘變粳稻品種春江06得到(輻射劑量為200Gy,劑量率為10Gy·h-1),表型上它與野生型相比有著多方面的改變。突變體mit1表現為分蘗中度增多、株高半矮的表型,成熟期株高稍矮,約為野生型的80%,各節間等比例一定程度的縮短;分蘗數目增多,約為野生型的2倍(見圖1);株型緊湊,分蘗角度和葉片角度小;插秧后的不同發育時期,mit1株高均低于野生型,分蘗均多于野生型(見圖2)。突變體mit1的育性結實正常,每穗粒數約為野生型70%,但單株總粒數及產量比野生型有所增加(見圖2)。
實施例2水稻MIT1基因的獲得和功能互補驗證
1、水稻MIT1基因的獲得
本發明的MIT1基因是采用圖位克隆方法通過突變體mit1克隆得到的。
經60Co-γ輻射誘變得到的突變體mit1經過多代自交種植確認該突變具有穩定遺傳的特性。純合突變體mit1與正常表型的秈稻Dular,進行雜交,所有雜交F1中個體均矮化分蘗中度增多表型。在F2種植群體中出現了明顯分離,通過調查分析,發現野生型與突變體的分離比例符合3:1遺傳分離比例。由此可以推測,該水稻矮化分蘗中度增多突變體的突變性狀是由1對隱性基因控制。
為了定位控制該分蘗中度增多突變體基因的位置,利用F1代自交構建的F2分離群體作為定位的群體,以BSA法選取l0株F2突變單株構建DNA混池,利用能較均勻覆蓋全基因組的170對Indel標記對兩個親本與基因池進行多態性篩選,發現與第12染色體上的R12-8和R12-10連鎖。為了精細定位目的基因,將F2定位群體擴大到1529株矮化分蘗中度增多個體,并開發了8對新的InDel標記,最終將目的基因定位在標記M4和M7之間(圖3的A圖)。這兩個標記之間的物理距離為2Mb,位于第12染色體的著絲粒區,包含15個候選基因。根據各基因預測的蛋白功能,首先選擇功能與本研究相關的6個基因進行測序,發現基因LOC_Os12g21710發生突變。
LOC_Os12g21710基因組DNA全長2937bp,包含4個外顯子和3個內含子,編碼區全長1104bp,共設計8對引物進行測序(引物序列見表1)。對基因測序結果分析發現,此基因在第二內含子和第三外顯子交界處發生10bp的缺失,包括第二內含子3’末端的2個堿基(ag)和第三外顯子5’端的8個堿基(GTCTACTT)(見圖3的B圖)。這10bp的缺失造成轉錄過程中第二內含子沒有被拼接掉而被錯誤讀取以及第三外顯子讀取的提前終止,從而造成基因的突變。將該基因命名為MIT1。根據水稻基因組注釋網站(http://rice.plantbiology.msu.edu/)預測,MIT1編碼15-順式-ζ-胡蘿卜素異構酶(15-cis-zeta-carotene isomerase,Z-ISO),它是類胡蘿卜素生物合成過程中一個重要的異構酶。
表1實施例1中涉及的引物序列
2、MIT1基因的功能互補驗證
本發明將MIT1基因克隆至植物表達載體pCAMBIA 1305.1(購自pCAMBIA公司),并在大腸桿菌TOP10中擴繁。通過農桿菌介導轉化方法,將pCAMBIA 1305.1攜帶的MIT1基因轉入水稻,獲得轉基因互補植株。
12g21710基因組互補載體構建:因為基因組DNA片段比較大,所以分成兩段擴增,以PstI位點為界限。第一個片段,5’端引入SacI位點,3’端引入PstI位點,片段長為4625bp,ATG前長度為1834bp,重組到pCAMBIA1305.1的SacI+PstI位點中去;第二個片段,兩端都引入PstI位點,片段長度為1240bp,TGA后長度為1086bp,重組到含第一個片段的載體的PstI位點中去。所用引物為12g21710S1SPF:ATGATTACGAATTCGAGCTCCAGTGGACAAAGCTTACCAAACTG和12g21710S1SPR:CCAAGCTTGCATGCCTGCAGCAAAAGGAAAACACTTG;12g21710S2PPF:TGTTATTCCTTTTGCTGCAGTTATCGATGGAAGAC和12g21710S2PPR:CCAAGCTTGCATGCCTGCAGCCTGGCACTTCTGATATAGGG。
互補植株的表型完全恢復正常,分蘗數和株高都和野生型一致。基因組DNA的互補結果證明了LOC_Os12g21710就是目的基因(見圖3的C圖)。
實施例3 MIT1的同源蛋白序列比對和進化分析
水稻Z-ISO(MIT1)、玉米Z-ISO(ZmZ-ISO)和擬南芥Z-ISO(AtZ-ISO)序列比對顯示同源性比較高。據水稻基因組注釋網站(http://rice.plantbiology.msu.edu/)預測,MTI1包含368個氨基酸,ChloroP 1.1Prediction Server預測該蛋白在N端有一個葉綠體轉運肽(方框部分),整個蛋白含有5個跨膜區(下劃線部分),還有一個NnrU結構域(見圖4的A圖)。
Z-ISO為15-順式-ζ-胡蘿卜素異構酶,是參與類胡蘿卜素生物合成途徑的關鍵酶之一。進化樹分析顯示,MTI1在高等植物和硅藻中都有同源基因,它們都有一個共同的起源—NnrU基因,該基因在反硝化細菌中發現,如根癌土壤桿菌(Agrobacterium fabrum str.C58)和苜蓿中華根瘤菌(Sinorhizobium meliloti 1021)(見圖4的B圖)。NnrU編碼一個跨膜蛋白—亞硝酸鹽和一氧化氮還原酶U(nitrite and nitric oxide reductase U,NnrU),最初在紅假單胞菌2.4.3中有描述,催化細菌反硝化作用。
實施例4水稻MIT1基因表達模式
提取粳稻品種日本晴不同組織(根、莖、葉片、葉鞘、側芽、花、穗)的RNA,反轉錄為cDNA進行Real-time PCR(RT-PCR)檢測,以水稻Ubiquitin基因為內參,檢測MIT1基因在水稻不同組織中的表達差異。
1、RNA提取
采用RNAprep pure Plant Kit(天根)進行水稻總RNA的提取。用于分析基因組織表達模式的樣品取材于抽穗期的野生型的根、莖、葉片、葉鞘、側芽、幼穗等部位。
(1)100mg樣品經液氮研磨后加入450μL裂解液,震蕩混勻。將溶液轉移至過濾柱中,12000rpm離心5min,收集管中上清至RNase-free的離心管中;
(2)加入1/2倍上清體積的無水乙醇,顛倒混勻后將溶液轉入吸附柱中,12000rpm離心1min,棄掉廢液;
(3)向吸附柱中加入350μL去蛋白液,12000rpm離心1min,棄掉廢液;
(4)向吸附柱中央加入80μL DNaseⅠ工作液,室溫放置15min;
(5)向吸附柱中加入350μL去蛋白液,12000rpm離心1min,棄掉廢液;
(6)向吸附柱中加入500μL漂洗液,室溫靜置2min,12000rpm離心1min,棄掉廢液;
(7)重復步驟(6),然后再12000rpm離心2min,棄掉廢液。將吸附柱置于一個新的RNase-free離心管中,室溫放置數分鐘;
(8)向吸附膜中央滴加30μL RNase-free ddH2O,室溫放置5min,12000rpm離心2min,RNA保存于-80℃;
2、cDNA合成
第一鏈cDNAs合成采用III First Strand Synthesis Kit(Invitrogen,USA),由于水稻基因GC含量較高,采用試劑盒推薦的高GC方法并且選用Oligo(dT)20(50μM)進行合成。具體操作方法如下:
(1)RNA/primer mixture的準備:1至5μg總RNA,1μL Oligo(dT)20(50μM),2.5μL dNTP mix(10mM),DEPC處理水補齊至25μL;
(2)將上述體系放入65℃孵環境下孵育5min后立即轉移至55℃;
(3)cDNA Synthesis Mix的準備(25μL):DEPC-treated water 3μL,10×RT buffer 5μL,10μL MgCl2(25mM),5μL DTT(0.1M),RNaseOUT Recombinant RNase Inhibitor 1μL,SuperScript III RT 1μL,上述混勻后置于55℃備用;
(4)將RNA/primer mixture與cDNA Synthesis Mix輕輕混勻,置于55℃,50min后轉入85℃,5min終止反應后冰上放置;
(5)最后加入1μL RNase H 37℃,20min,-20℃存儲備用。
3、實時定量RT-PCR
Real-time PCR實驗采用Premix Ex TaqTM II(Perfect Real Time)試劑盒(寶生物),在Applied Biosystems公司的7300HT Real-Time PCR儀上進行。以水稻ubiquitin基因作為內參。每個反應做三次生物學重復,相對表達量計算方法參見2-△△CT法。Real-time PCR程序:95℃30s;95℃5s,60℃30s,共40個循環;95℃15s,60℃1min,95℃15s。
Real-time PCR反應體系如下:
結果表明,MIT1在葉鞘中表達量高,其次是側芽、根和花,在莖和穗中表達量較低(見圖5)。
實施例5水稻組織GUS染色結果
12g21710Prom::GUS載體構建:MIT1啟動子區通過PCR擴增獲得,在5’端引入BamHI位點,3’端引入NcoI位點,片段長為2460bp,為翻譯水平融合,包含前7個氨基酸在內。重組到pCAMBIA1305.1的BamHI和NcoI位點中去。所用引物為12g21710PromBNF:CGGTACCCGGGGATCCGGAGAACCTGTCTGCACATTC,12g21710Prom BNG:CTCAGATCTACCATGGGGAGGA GAGATGGGAGGCCAT。利用農桿菌侵染水稻愈傷的方法獲得轉基因植株,用于分析基因組織表達模式。
利用GUS(β-glucuronidase)基因作為報告基因,檢測了MIT1基因表達的空間分布情況。MIT1表達模式與RT-PCR結果一致,在莖、節、根、穎殼、葉片和葉鞘中均能檢測到GUS活性。在莖、穎殼和葉鞘中GUS活性較強(見圖6)。
實施例6 MIT1蛋白的亞細胞定位
35S::MIT1-GFP載體構建:以水稻日本晴cDNA為模板擴增MIT1的全長編碼區,片段長1104bp,片段5'和3'端分別引入酶切位點。用相應的高保真內切酶酶切空載體pAN580成線性載體,應用In-Fusion高保真克隆試劑盒說明書連接載體和片段,熱激法轉化大腸桿菌TOP10感受態細胞。PCR檢測獲得陽性克隆,菌液送公司測序正確后備用。所用引物為:12g21710GFPF:AGCCCAGATCAACTAGTATGGCCTCCCATCTCCTCCTC,12g21710GFPR:TTGCTCACCATGGATCCCCAAGGGAGTTGGTAGCTGGA。
瓊脂糖凝膠檢測,膠回收試劑盒(GenStar)回收目的片段;
(1)獲得目的片段:PCR擴增
(2)載體準備:所使用的內切酶是NEB的。
37℃,酶切3h。瓊脂糖凝膠檢測,膠回收試劑盒(GenStar)回收目標載體。
50℃連接,15min。冰上放置2min。向其中加入50μL的感受態細胞,比那個上放置30min.
(4)轉化:將上述混好的體系,42℃,熱擊45s。冰上放置2min。
加入無抗生素的LB培養基,37℃,搖培45min-1h。
(5)涂板,平板倒置,37℃培養14-16h。
(6)挑單克隆,測序檢測正確的保菌保存,提質粒保存。
為了闡述MIT1的亞細胞定位情況,利用綠色熒光蛋白(GFP)作為標簽,構建了35S::MIT1-GFP載體,將其轉入到野生型的原生質體內,觀察融合蛋白MIT1:GFP的瞬時表達,并以35S::GFP的瞬時表達作為對照。結果顯示由35S啟動子啟動的GFP蛋白在在細胞膜、細胞質和細胞核內均有表達,而MIT1:GFP蛋白只在葉綠體里表達(見圖7)。所以,蛋白MIT1定位在葉綠體上。
以上所述僅是本發明的優選實施方式,應當指出,對于本技術領域的普通技術人員來說,在不脫離本發明技術原理的前提下,還可以做出若干改進和潤飾,這些改進和潤飾也應視為本發明的保護范圍。
- 還沒有人留言評論。精彩留言會獲得點贊!
- 蝎神經毒素AaIT及其編碼基因與應用的制造方法與工藝
- 腈水解酶突變體及其編碼基因和應用的制造方法與工藝
- 植物抗旱相關蛋白AsTMK1及其編碼基因和應用的制造方法與工藝
- 植物抗旱相關蛋白EtSnRK2.2及其編碼基因和應用的制造方法與工藝
- OsSAPK7蛋白及其編碼基因在提高水稻白葉枯病抗性中的應用的制造方法與工藝
- 一種抗除草劑抗蟲融合基因、編碼蛋白及應用的制造方法與工藝
- 毛果楊PtrZFP103基因及其編碼蛋白和應用的制造方法與工藝
- 大豆bHLH轉錄因子基因GmFER及其編碼蛋白和應用的制造方法與工藝
- 一種花狹口蛙的基因及其編碼的多肽和該多肽的用途的制造方法與工藝
- 一種澤陸蛙基因及其編碼的多肽和該多肽的用途的制造方法與工藝
- 降低種子重量的白樺AP2基因及其編碼蛋白的制造方法與工藝
- 可編碼ArtRD蛋白基因及其蛋白和應用的制造方法與工藝
- 蝎神經毒素AaIT及其編碼基因與應用的制造方法與工藝
- 腈水解酶突變體及其編碼基因和應用的制造方法與工藝
- 植物抗旱相關蛋白AsTMK1及其編碼基因和應用的制造方法與工藝
- 植物抗旱相關蛋白EtSnRK2.2及其編碼基因和應用的制造方法與工藝
- OsSAPK7蛋白及其編碼基因在提高水稻白葉枯病抗性中的應用的制造方法與工藝
- 一種抗除草劑抗蟲融合基因、編碼蛋白及應用的制造方法與工藝
- 毛果楊PtrZFP103基因及其編碼蛋白和應用的制造方法與工藝
- 大豆bHLH轉錄因子基因GmFER及其編碼蛋白和應用的制造方法與工藝
- 腈水解酶突變體及其編碼基因和應用的制造方法與工藝
- 植物抗旱相關蛋白AsTMK1及其編碼基因和應用的制造方法與工藝
- 植物抗旱相關蛋白EtSnRK2.2及其編碼基因和應用的制造方法與工藝
- OsSAPK7蛋白及其編碼基因在提高水稻白葉枯病抗性中的應用的制造方法與工藝
- 一種抗除草劑抗蟲融合基因、編碼蛋白及應用的制造方法與工藝
- 毛果楊PtrZFP103基因及其編碼蛋白和應用的制造方法與工藝
- 大豆bHLH轉錄因子基因GmFER及其編碼蛋白和應用的制造方法與工藝
- 一種花狹口蛙的基因及其編碼的多肽和該多肽的用途的制造方法與工藝
- 一種澤陸蛙基因及其編碼的多肽和該多肽的用途的制造方法與工藝
- 一種水稻MIT1基因、其編碼蛋白及應用的制造方法與工藝
- 植物抗旱相關蛋白EtSnRK2.2及其編碼基因和應用的制造方法與工藝
- OsSAPK7蛋白及其編碼基因在提高水稻白葉枯病抗性中的應用的制造方法與工藝
- 一種抗除草劑抗蟲融合基因、編碼蛋白及應用的制造方法與工藝
- 毛果楊PtrZFP103基因及其編碼蛋白和應用的制造方法與工藝
- 大豆bHLH轉錄因子基因GmFER及其編碼蛋白和應用的制造方法與工藝
- 一種水稻MIT1基因、其編碼蛋白及應用的制造方法與工藝
- 蛋白StTrxF、編碼基因及其在提高植物淀粉含量中的應用的制造方法與工藝
- 蛋白質及其編碼基因在調控植物抗黃萎病性中的應用的制造方法與工藝
- 甘薯耐逆相關蛋白IbFBA2、編碼基因、重組載體與應用的制造方法與工藝
- 植物抗旱相關蛋白ZmNAC111及其編碼基因與應用的制造方法與工藝
- 植物抗旱相關蛋白EtSnRK2.2及其編碼基因和應用的制造方法與工藝
- OsSAPK7蛋白及其編碼基因在提高水稻白葉枯病抗性中的應用的制造方法與工藝
- 一種抗除草劑抗蟲融合基因、編碼蛋白及應用的制造方法與工藝
- 毛果楊PtrZFP103基因及其編碼蛋白和應用的制造方法與工藝
- 大豆bHLH轉錄因子基因GmFER及其編碼蛋白和應用的制造方法與工藝
- 一種水稻MIT1基因、其編碼蛋白及應用的制造方法與工藝
- 蛋白StTrxF、編碼基因及其在提高植物淀粉含量中的應用的制造方法與工藝
- 植物抗旱相關蛋白ZmNAC111及其編碼基因與應用的制造方法與工藝
- 核盤菌凝集素SSL?6蛋白及其編碼基因與應用的制造方法與工藝
- 植物抗病毒相關蛋白GmSN1及其編碼基因和應用的制造方法與工藝
- 植物抗旱相關蛋白EtSnRK2.2及其編碼基因和應用的制造方法與工藝
- OsSAPK7蛋白及其編碼基因在提高水稻白葉枯病抗性中的應用的制造方法與工藝
- 一種抗除草劑抗蟲融合基因、編碼蛋白及應用的制造方法與工藝
- 毛果楊PtrZFP103基因及其編碼蛋白和應用的制造方法與工藝
- 大豆bHLH轉錄因子基因GmFER及其編碼蛋白和應用的制造方法與工藝
- 一種水稻MIT1基因、其編碼蛋白及應用的制造方法與工藝
- 蛋白StTrxF、編碼基因及其在提高植物淀粉含量中的應用的制造方法與工藝
- 蛋白質及其編碼基因在調控植物抗黃萎病性中的應用的制造方法與工藝
- 植物抗旱相關蛋白ZmNAC111及其編碼基因與應用的制造方法與工藝
- 核盤菌凝集素SSL?6蛋白及其編碼基因與應用的制造方法與工藝