一種降龍涎醚的生物催化合成方法
本發明屬于生物催化和合成生物學領域,提供了一種化學-酶聯用的合成方法,從生物合成來源的萜類化合物如橙花叔醇開始,將簡潔的化學轉化與下游的生物酶轉化法相結合,用于高效合成香料成分降龍涎醚(ambroxide)。
背景技術:
1、天然龍涎香是一種名貴香料,它來自抹香鯨腸胃中的灰白色軟結石,因量極少而價比黃金。由于天然龍涎香的來源稀缺且獲取困難,相關研究方向多致力于開發人工合成方法,以替代天然資源。降龍涎醚[(-)-ambroxide]是天然龍涎香最關鍵的有效成分之一,具有類似木質、琥珀和煙熏的香味(l.cristiano等人,cosmetics?2022,9,13)。降龍涎醚在香料和化妝品工業中具有重要價值,是所有合成龍涎香香料中評價最高的一種,因其獨特的香氣、穩定性及良好的揮發性,可增加香料的持久性和擴散性,作為龍涎香優異代用品之一和極其優秀的定香劑,被廣泛應用于高級香水和香精的生產中。由于同時降龍涎醚對人體無刺激、無過敏反應,因此很適合用于皮膚、頭發和織物的加香,在化妝品行業中也被廣泛應用(n.he等人,chem&bio?engineering,2024,1(2):91)。同時降龍涎醚也應用于卷煙工業的加香加料,改善煙草的香味品質(n.he等人,acs?sustain?chem?eng,2023,11:1939)。
2、現有降龍涎醚主要的獲取方法如下所示:
3、
4、降龍涎醚骨架為16個碳原子的類半日花烷型萜類,屬于含有非經典碳原子數量的萜烯衍生化合物。目前降龍涎醚的工業生產主要以香紫蘇醇(sclareol)、β-二氫紫羅蘭酮、金合歡醇類似物為原料進行制備,其中香紫蘇醇具有和降龍涎醚相似的碳原子骨架,可以合成得到光學純降龍涎醚,是合成降龍涎醚最為實用的原料,該路線也是目前產業界獲取降龍涎醚主要方法。具體地,該路線以植物提取的天然產物香紫蘇醇為原料,進行化學氧化、還原、環化反應,先后形成香紫蘇內酯和香紫蘇二醇,最終產生降龍涎醚。但這些生產方法從綠色化學,立體化學和供應鏈的角度來看依然存在諸多的問題。如香紫蘇醇技術路線的關鍵步驟是用對側鏈進行氧化降解得到香紫蘇內酯,須用到等化學當量的污染較為嚴重的氧化劑。例如,傳統的合成方法是鉻酸酐氧化法,即用鉻酸酐氧化香紫蘇醇來合成,操作容易,但在生產過程中產生的含鉻廢水對環境污染較大,現在基本不再采用。barton等人提出用高碘酸鈉和四氧化鋨在四氫呋喃中氧化香紫蘇醇。隨后美國reynolds公司開發了高錳酸鉀兩步氧化法(us3050532)。中國上海香料所也開發出了高錳酸鉀低溫氧化工藝等。這些氧化劑可對香紫蘇醇側鏈進行氧化降解而獲得香紫蘇內酯,但均會造成嚴重的重金屬污染。盡管使用綠色氧化劑臭氧、雙氧水等進行氧化降解的方法已有報道,但其收率和產品分離效率尚不能滿足工業化需求,且化學試劑的殘留以及立體選擇性較差等原因極大的限制了這些合成方法的應用。此外這些半合成路線的成本受植物提取的香紫蘇醇供應情況波動較大,不利于產品成本的控制。最后由于降龍涎醚具有多個不同的立體異構體結構,異構體之間具有一定的香氣特征差異,化學合成中引入的各類降龍涎醚類似物和光學異構體,影響了終產品的純度和香型。
5、目前天然提取物仍然是天然香料香精市場的主要來源,但是面臨全球人口不斷增長和耕地面積日趨減少的壓力,對于解決人類溫飽問題的農作物的需求顯然比香料作物更為迫切。從長遠發展來看,尋找可替代香料作物的天然香料來源成為一種必然的趨勢,而生物技術制備的香料被視為天然產品,在消費端市場具有一定的溢價。此外,與傳統的有機合成工藝相比,生物技術還具有反應條件溫和、立體選擇性好、環境污染小等優點,因此利用生物技術制備天然香料成為中香料領域最受關注的研究熱點之一。生物轉化和化學催化相結合的方式生產降龍涎醚已有報道:一種是利用真菌cryptococcus?albidus?atcc?20918或filobasidium?magnum?jd1025以香紫蘇醇為原料發酵生產香紫蘇內脂(us5212078a,1993)再經化學轉化合成降龍涎醚;另一種方式是用真菌hyphozyma?roseoniger?atcc20624以化合物為原料生產香紫蘇二醇(us4798799a,1989),再以香紫蘇二醇為原料通過化學法生產降龍涎醚。紅絲酵母(hyphozyma?roseonigra?atcc?20624)是目前已知的唯一可以利用香紫蘇醇為單一碳源,以高轉化率代謝生成香紫蘇二醇的微生物。從香紫蘇醇出發由紅絲酵母直接轉化生成香紫蘇二醇,再經一步簡單的脫水環化得到降龍涎醚是一種有吸引力的替代方法,然而香紫蘇醇對紅絲酵母具有細胞毒性,因此經濟上可行的底物濃度難以達到。此外其代謝通路一直不明了,阻礙了我們在其他底盤細胞中異源構建降龍涎醚高產菌株或高效生物催化體系。已有研究提出香紫蘇醇可能經過紅絲酵母中未知醇醛脫氫酶的轉化生成相應的α,β-不飽和酸,但是其后續的代謝產物和代謝酶仍然需要進一步解析。
6、美國奇華頓公司(givaudan)研發了一條從β-法尼烯起始,經由ee-高法尼醇,酶催化法一步合成光學純降龍涎醚的技術路線ambrofix(wo2016/170099,wo2021/110848)。與從香紫蘇醇出發的半化學合成路線相比,ambrofix把多步收率低污染重的化學合成轉變為一步高效角鯊烯何帕烯環化酶(shc酶)催化環化,實現了最大程度的原子經濟性和綠色可持續性,是降龍涎醚生物合成領域近年來最大的創新。但是在該路線中,ee-高法尼醇仍然需要多步的危險性較高、污染較重的化學合成步驟來獲取,且該路線因為底物在化學轉化過程中發生的順反異構推高了ee-高法尼醇合成成本。該路線始于生物基(e)-構象的β-法尼烯,由甘蔗為碳源發酵,由釀酒酵母異源表達法尼烯合成酶生產(wo2015/059293)。在化學轉化中,首先在鈀(ii)催化劑存在下,由n-亞硝基-n-甲基脲(nmu)和氫氧化鉀原位生成重氮甲烷,對(e)-構象的β-法尼烯端位雙鍵進行選擇性環丙化。雖然這個過程是在流動系統中連續進行,原子經濟較高,但nmu是一種強致癌物、誘變劑和致畸劑,因此該過程仍然不符合綠色化學的要求。環丙化完成后,通過相轉移介導的酸化反應進行環丙基重排,并轉化為相應的酯,脫酰化后生成ee-高法尼醇,該過程易產生順反異構副產物,不僅推高了合成成本,也影響了終產品的香型和光學純度。
7、因此,仍然需要利用合成生物學和生物催化技術,找到一條有市場競爭力的并且滿足綠色化學要求的降龍涎醚合成途徑,使其能夠滿足消費者對于天然香氛不斷增長的需求。
8、現有降龍涎醚及其類似物化學合成和生物合成方法存在如下缺陷:
9、1.現有降龍涎醚的化學合成路線須用到污染較為嚴重的重金屬氧化劑如四氧化鋨,高錳酸鉀等,收率低,分離純化成本高,同時反應原料香紫蘇醇來源于植物提取,成本受天氣變化影響波動高,供應鏈穩定性差。
10、2.現有生物轉化獲取降龍涎醚仍然需要下游的化學轉化,且底物與純化學合成路線相同,均為植物提取來源的香紫蘇醇,仍然面臨供應鏈波動問題。此外底物香紫蘇醇天然對細胞有毒性,生物轉化的底物濃度尚不能滿足工業化高效生產的需要。
11、3.現有生物催化路線如ambrofix仍然需要使用高毒性的試劑來合成酶催化所需底物,不能滿足綠色化學的要求,且化學合成中引入的底物順反異構推高了該路線的生產成本。
技術實現思路
1、為了解決現有技術中的問題,第一方面本發明提供一種拜爾-維利格單加氧酶及其天然共表達的酯酶和醇脫氫酶、拜爾-維利格單加氧酶與醇脫氫酶融合蛋白、拜爾-維利格單加氧酶與酯酶的融合蛋白、角鯊烯何帕烯環化酶突變體的氨基酸序列。
2、第二方面本發明提供一種編碼基因,編碼本發明第一方面所述拜爾-維利格單加氧酶、天然共表達的酯酶、醇脫氫酶、角鯊烯何帕烯環化酶突變體的基因、以及所述關鍵酶融合蛋白的基因。
3、第三方面本發明提供含有本發明第二方面所述編碼拜爾-維利格單加氧酶、酯酶、醇脫氫酶、角鯊烯何帕烯環化酶突變體、關鍵酶融合蛋白的編碼基因的重組質粒。
4、第四方面本發明提供了含有本發明第三方面所述的重組質粒的基因工程菌及其在生物催化法合成降龍涎醚中的應用。
5、第五方面本發明提供一種降龍涎醚的生物催化合成方法,以克服現有技術中反應原料香紫蘇醇來源于植物提取,供應鏈穩定性差,化學轉化重金屬污染重;底物香紫蘇醇天然對細胞有毒性,生物轉化的底物濃度尚不能滿足工業化高效生產的需要;生物催化路線需要使用高毒性的試劑的不足。
6、為實現前述發明目的,本發明采用的技術方案包括:
7、一種拜爾-維利格單加氧酶及其天然共表達的酯酶和醇脫氫酶,所述拜爾-維利格單加氧酶的氨基酸序列為seq?id?no:3所示,所述酯酶的氨基酸序列為seq?id?no:4-6所示,所述醇脫氫酶的氨基酸序列為seq?id?no:7所示。
8、一種融合蛋白,使用柔性連接子對的拜爾-維利格單加氧酶與醇脫氫酶、或拜爾-維利格單加氧酶與酯酶進行融合,所述拜爾-維利格單加氧酶的氨基酸序列選自seq?idno:1-3其中一種,所述酯酶的氨基酸序列選自seq?id?no:4-6其中一種,所述醇脫氫酶的氨基酸序列為seq?id?no:7所示。
9、所述拜爾-維利格單加氧酶與醇脫氫酶融合蛋白的氨基酸序列如seq?id?no:18-19所示,所述拜爾-維利格單加氧酶與酯酶的融合蛋白氨基酸序列如seq?id?no:20-21所示。
10、除所列出的氨基酸序列外,可以使用本領域已知的任何合適的形式柔性連接子對拜爾-維利格單加氧酶與醇脫氫酶、或拜爾-維利格單加氧酶與酯酶進行融合。例如但不限于:gsg,ggggs,(ggggs)2,(ggggs)3,eaaak,(eaaak)2,(eaaak)3,g8,g6。
11、一種將法尼基丙酮轉化為ee-高法尼醇的方法,包括如下步驟:
12、步驟1在輔因子存在下拜爾-維利格單加氧酶催化法尼基丙酮轉化為高法尼醇乙酸酯,所述拜爾-維利格單加氧酶的氨基酸序列選自seq?id?no:1-3其中一種;
13、步驟2酯酶催化高法尼醇乙酸酯脫乙酰化得到ee-高法尼醇,酯酶的氨基酸序列選自
14、seq?id?no:4-6其中一種。
15、優選地,所述拜爾-維利格單加氧酶、酯酶的具體形式為大腸桿菌粗提物上清。所述拜爾-維利格單加氧酶、酯酶、醇脫氫酶、角鯊烯何帕烯環化酶突變體和關鍵酶融合蛋白可作為從相應的產生它的細胞制備的細胞提取物存在于反應混合物中。同理,也可用單一的細胞株表達所上述拜爾-維利格單加氧酶、酯酶、醇脫氫酶、角鯊烯何帕烯環化酶突變體和關鍵酶融合蛋白,其細胞提取物可以作為單一的生物催化劑用于體外生物催化合成降龍涎醚。優選地,所述細胞優選是用于產生所述酶的細菌宿主細胞。
16、細胞提取物可以通過能夠裂解宿主細胞的任何合適的方法獲得,包括但不限于:超聲處理、rna酶/溶菌酶處理、凍融處理或堿處理。所述細胞裂解物可以通過本領域已知的任何合適的方法處理,包括但不限于:過濾、離心或用鹽純化以獲得澄清的細胞提取物。
17、優選地,所述輔因子選自nadh和nadph或其組合。
18、所述方法包括輔因子再生系統。
19、優選地,所述輔因子再生系統為醇脫氫酶/異丙醇輔因子或葡萄糖脫氫酶(gdh)/葡萄糖再生系統。
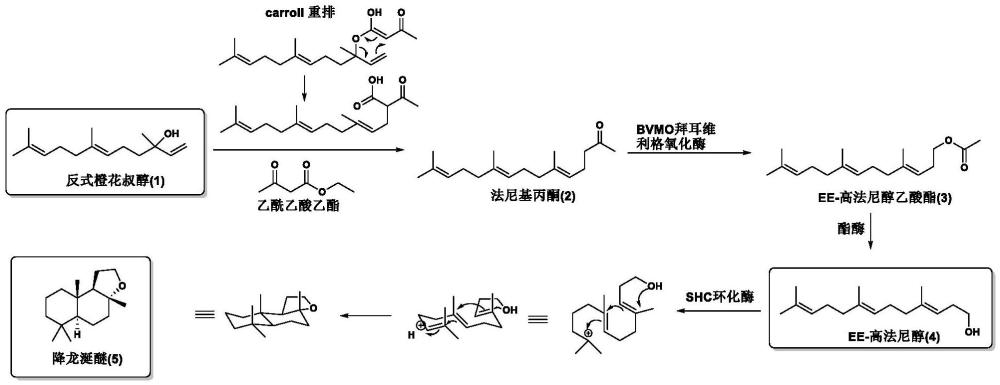
20、一種將法尼基丙酮轉化為ee-高法尼醇的方法,包括如下步驟:在輔因子存在下,使用選自拜爾-維利格單加氧酶與酯酶的融合蛋白其中一種,催化法尼基丙酮轉化為ee-高法尼醇。
21、一種將法尼基丙酮轉化為降龍涎醚的方法,包括如下步驟:
22、步驟1在輔因子存在下拜爾-維利格單加氧酶催化法尼基丙酮轉化為高法尼醇乙酸酯,所述拜爾-維利格單加氧酶的氨基酸序列選自seq?id?no:1-3其中一種;
23、步驟2酯酶催化高法尼醇乙酸酯脫乙酰化得到ee-高法尼醇,酯酶的氨基酸序列選自seq?id?no:4-6其中一種;
24、步驟3使用角鯊烯何帕烯環化酶突變體將ee-高法尼醇酶促轉化為降龍涎醚,所述角鯊烯何帕烯環化酶突變體的氨基酸序列選自seq?id?no:8-17其中一種。
25、反應混合物的ph可以在4至9,優選地在7至9的范圍內(其包括約8.5的ph),并且可以通過向反應混合物中添加緩沖液或ph校正劑來維持。用于此目的的示例性緩沖液為tris-hcl緩沖液或甘氨酸/naoh緩沖液。對于所考慮的bvmo酶,溫度為約15℃至約60℃。在生物轉化過程中,溫度可以保持恒定或者可以被改變。
26、一種編碼基因,所述基因編碼所述的拜爾-維利格單加氧酶的基因,所述編碼基因的核苷酸序列如seq?id?no:22-24所示;所述基因編碼所述的酯酶的基因,所述編碼基因的核苷酸序列如seq?id?no:25-27所示;所述基因編碼所述的醇脫氫酶的基因,所述編碼基因的核苷酸序列如seq?id?no:28所示。所述基因編碼所述的角鯊烯何帕烯環化酶突變體的基因,所述編碼基因的核苷酸序列如seq?id?no:29-38所示;所述基因編碼所述的拜爾-維利格單加氧酶與共表達的醇脫氫酶融合蛋白的基因,所述編碼基因的核苷酸序列如seq?idno:39-40所示;所述基因編碼所述的拜爾-維利格單加氧酶與酯酶的融合蛋白的基因,所述編碼基因的核苷酸序列如seq?id?no:41-42所示。
27、一種重組質粒,包含所述編碼拜爾-維利格單加氧酶的基因,所述重組質粒的核苷酸序列如seq?id?no:43-45所示;包含所述編碼酯酶的基因,所述重組質粒的核苷酸序列如seq?id?no:46-48所示;包含所述編碼醇脫氫酶的基因,所述重組質粒的核苷酸序列如seqid?no:49所示;包含所述編碼角鯊烯何帕烯環化酶突變體的基因,所述重組質粒的核苷酸序列如seq?id?no:50-59所示;包含所述編碼拜爾-維利格單加氧酶與共表達的醇脫氫酶融合蛋白的基因,所述重組質粒的核苷酸序列如seq?id?no:60-61所示;包含所述編碼拜爾-維利格單加氧酶與酯酶的融合蛋白的基因,所述重組質粒的核苷酸序列如seq?id?no:62-63所示。
28、所述重組質粒的表達載體為pet28a(+)。
29、一種基因工程菌,所述基因工程菌包含所述的重組質粒。
30、所述基因工程菌的表達宿主為e.coli?bl21(de3)。
31、所述基因工程菌在轉化線性多不飽和支鏈酮底物至相應的乙酸酯和碳鏈縮短兩個碳原子的醇,利用角鯊烯何帕烯環化酶進行酶促環化,合成含有非經典碳原子數量的萜烯衍生化合物中的應用。
32、線性多不飽和支鏈酮為法尼基丙酮,含有非經典碳原子數量的萜烯衍生化合物為降龍涎醚。
33、一種將橙花叔醇轉化為降龍涎醚的方法,包括如下步驟:
34、步驟1從反式橙花叔醇開始,進行carroll重排反應合成法尼基丙酮,橙花叔醇精餾與化學合成法尼基丙酮耦合;
35、步驟2在輔因子存在下拜爾-維利格單加氧酶催化法尼基丙酮轉化為高法尼醇乙酸酯,所述拜爾-維利格單加氧酶的氨基酸序列選自seq?id?no:1-3其中一種;
36、步驟3酯酶催化高法尼醇乙酸酯脫乙酰化得到ee-高法尼醇,酯酶的氨基酸序列選自seq?id?no:4-6其中一種;
37、步驟4使用角鯊烯何帕烯環化酶突變體將ee-高法尼醇酶促轉化為降龍涎醚,所述角鯊烯何帕烯環化酶突變體的氨基酸序列選自seq?id?no:8-17其中一種。
38、與現有技術相比,本發明的有益效果在于:
39、1)本發明中所用化學轉化部分的方法避免了使用任何高污染試劑,完全使用成本低廉,對操作人員和環境友好的無毒試劑。
40、2)同時技術路線優化了反應過程,將橙花叔醇的分離與化學轉化進行耦合,提高了生產效率,降低了能耗。
41、3)本發明采用了酶粗提物體外生物催化法,無活細胞參與轉化,因此繞過了因底物對細胞的抑制作用造成的單位體積下轉化濃度限制,提高了工業規模下該路線的實用性。
42、4)本發明采用了全新的酶催化法來獲取環化反應的關鍵底物ee-高法尼醇,轉化過程安全無毒,滿足了綠色化學的要求。同時我們使用了與關鍵酶新的拜爾-維利格單加氧酶、天然共表達的酯酶、醇脫氫酶以及與該系統相匹配的角鯊烯何帕烯環化酶突變體協同轉化,不僅促使氧化反應的輔酶循環使用而無需外源添加昂貴的輔酶,還可高效專一地催化從底物法尼基丙酮開始到降龍涎醚的轉化,而無需進行中間體的分離純化即可從反應體系中直接獲取終產品。
43、5)本發明所發現的bvmo酶活性相比現有解決方案更好。
44、6)本發明的shc突變體經過專門的進化和篩選,滿足了在復雜催化體系中要求的高適配性、穩定性和催化效率。
- 還沒有人留言評論。精彩留言會獲得點贊!