一種應(yīng)用磷脂酶C將胞內(nèi)蛋白在胞外表達(dá)的方法與流程
本發(fā)明涉及一種應(yīng)用磷脂酶C將胞內(nèi)蛋白在胞外表達(dá)的方法,屬于生物工程技術(shù)領(lǐng)域。
背景技術(shù):
大腸桿菌(Escherichia coli)表達(dá)系統(tǒng)作為目前應(yīng)用最廣泛、研究最透徹的表達(dá)系統(tǒng)之一,在培養(yǎng)周期長(zhǎng)短、實(shí)驗(yàn)操作簡(jiǎn)便度等方面具有顯著優(yōu)勢(shì)。由于大腸桿菌具有兩層的細(xì)胞膜結(jié)構(gòu),外源基因在大腸桿菌表達(dá)系統(tǒng)中的表達(dá)產(chǎn)物可能定位于大腸桿菌空間結(jié)構(gòu)的不同位置,通常有三種定位方式:胞內(nèi)、胞外和周質(zhì)空間定位。
有研究發(fā)現(xiàn),當(dāng)天然胞內(nèi)定位的蛋白在大腸桿菌表達(dá)系統(tǒng)重組表達(dá)時(shí),即使給目的蛋白添加信號(hào)肽,也不能穿過(guò)大腸桿菌的細(xì)胞膜分泌到細(xì)胞外。這些蛋白通常只能在細(xì)胞內(nèi)進(jìn)行表達(dá)并且需要利用物理或化學(xué)方法對(duì)細(xì)胞進(jìn)行破碎才能獲得目的蛋白,后續(xù)提取工藝不但繁瑣而且成本較高,因此尋找能夠有效的簡(jiǎn)化下游提取步驟降低產(chǎn)物純化的成本成為了研究者的目標(biāo)。
技術(shù)實(shí)現(xiàn)要素:
本發(fā)明的第一個(gè)目的是提供一種基因工程菌,是以大腸桿菌為宿主,表達(dá)磷脂酶C;所述磷脂酶C的氨基酸序列如SEQ ID NO.1-3任一所示。
在本發(fā)明的一種實(shí)施方式中,所述基因工程菌共表達(dá)磷脂酶C與目標(biāo)蛋白;所述目標(biāo)蛋白是胞內(nèi)蛋白,該基因工程菌能夠使胞內(nèi)蛋白在胞外表達(dá)。
在發(fā)明的一種實(shí)施方式中,三種磷脂酶C基因分別來(lái)源于Listeria monocytogenes、Clostridium perfringens和Bacillus cereus。
在本發(fā)明的一種實(shí)施方式中,所述目標(biāo)蛋白是胞內(nèi)定位蛋白;所述胞內(nèi)定位蛋白是在伴侶蛋白的輔助下,合成于核糖體后仍定位在細(xì)胞基質(zhì)中的蛋白。
在本發(fā)明的一種實(shí)施方式中,所述目標(biāo)蛋白包括蔗糖磷酸化酶,海藻糖酶。
在本發(fā)明的一種實(shí)施方式中,所述磷脂酶C與目標(biāo)蛋白是以pETDuet-1為載體進(jìn)行共表達(dá)。
在本發(fā)明的一種實(shí)施方式中,所述基因工程菌以E.coli BL21(DE3)為宿主。
本發(fā)明的第二個(gè)目的是提供所述基因工程菌的構(gòu)建方法,是將編碼磷脂酶C的基因和編碼目標(biāo)蛋白的基因分別連接到pETDuet-1表達(dá)載體的多克隆位點(diǎn)MCS1或MCS2上,得到重組表達(dá)質(zhì)粒,然后將重組質(zhì)粒轉(zhuǎn)化到宿主菌中。
在本發(fā)明的一種實(shí)施方式中,所述編碼磷脂酶C的基因包括SEQ ID NO.4-6。
在本發(fā)明的一種實(shí)施方式中,所述宿主菌為E.coli BL21(DE3)。
本發(fā)明的第三個(gè)目的是提供一種將胞內(nèi)蛋白在胞外表達(dá)的方法,是將磷脂酶C分別與目標(biāo)蛋白在大腸桿菌中共表達(dá);所述磷脂酶C的氨基酸序列分別為SEQ ID NO.1、SEQ ID NO.2和SEQ ID NO.3所示。
本發(fā)明的第四個(gè)目的是提供一種生產(chǎn)胞外蛋白的方法,所述方法是將所述基因工程菌接種至發(fā)酵培養(yǎng)基中進(jìn)行發(fā)酵,離心,收集發(fā)酵上清液。
在本發(fā)明的一種實(shí)施方式中,具體是:(1)分別將基因序列如SEQ ID NO.4-6所示的三種磷脂酶C基因和蔗糖磷酸化酶基因連接到pETDuet-1表達(dá)載體的多克隆位點(diǎn)MCS1或MCS2上,得到重組表達(dá)質(zhì)粒,(2)將重組質(zhì)粒轉(zhuǎn)化到宿主菌,(3)對(duì)重組菌進(jìn)行發(fā)酵培養(yǎng),收集發(fā)酵上清液。
在本發(fā)明的一種實(shí)施方式中,所述表達(dá)載體為pETDuet-1,所述宿主菌為E.coli BL21(DE3)。
在本發(fā)明的一種實(shí)施方式中,所述目標(biāo)蛋白為胞內(nèi)定位蛋白;所述胞內(nèi)定位蛋白是在伴侶蛋白的輔助下,合成于核糖體后仍定位在細(xì)胞基質(zhì)中的蛋白。
在本發(fā)明的一種實(shí)施方式中,所述目標(biāo)蛋白包括蔗糖磷酸化酶,海藻糖酶。
在本發(fā)明的一種實(shí)施方式中,所述方法具體包括如下步驟:
(1)將SEQ ID NO.4-6所示不含信號(hào)肽的磷脂酶C成熟蛋白基因以及蔗糖磷酸化酶基因,連接至pMD19-T simple vector,獲得質(zhì)粒pMD19T-plc(以plc代表三種磷脂酶C)和pMD19T-sp;
(2)分別對(duì)表達(dá)載體pETDuet-1和上步構(gòu)建成功的pMD19T-plc或pMD19T-sp質(zhì)粒進(jìn)行雙酶切,將不同的目的基因分別連接到表達(dá)載體pETDuet-1的多克隆位點(diǎn)MCS1,轉(zhuǎn)化E.coli JM109感受態(tài)細(xì)胞,得到重組質(zhì)粒pETDuet-1-plc和pETDuet-1-sp。
(3)分別對(duì)含有sp和plc基因的質(zhì)粒以及上步構(gòu)建的pETDuet-1-plc和pETDuet-1-sp進(jìn)行雙酶切,將sp或plc分別連接到重組質(zhì)粒pETDuet-1-plc和pETDuet-1-sp的多克隆位點(diǎn)MCS2,轉(zhuǎn)化E.coli JM109感受態(tài)細(xì)胞,得到共表達(dá)質(zhì)粒pETDuet-1-plc-sp和pETDuet-1-sp-plc。
(4)將步驟(3)構(gòu)建的共表達(dá)質(zhì)粒pETDuet-1-plc-sp和pETDuet-1-sp-plc分別導(dǎo)入宿主大腸桿菌,獲得六株重組菌;
(5)發(fā)酵培養(yǎng)步驟(4)構(gòu)建的重組大腸桿菌生產(chǎn)目標(biāo)蛋白,收集發(fā)酵上清液。
在本發(fā)明的一種實(shí)施方式中,所述發(fā)酵培養(yǎng)具體是:將種子以8-10%的接種量接種,誘導(dǎo)前培養(yǎng)溫度為37℃,誘導(dǎo)后培養(yǎng)溫度30℃,pH6.5-7.5,溶氧20-30%;當(dāng)發(fā)酵初始碳源甘油基本消耗完時(shí),開(kāi)始流加補(bǔ)料液,所述補(bǔ)料液按指數(shù)流加,比生長(zhǎng)速率控制在μ=0.1-0.3h-1;當(dāng)菌體細(xì)胞濃度達(dá)到OD600為30-50時(shí),開(kāi)始誘導(dǎo),誘導(dǎo)方法為恒速流加乳糖,乳糖加量0.1-0.4g·L-1·h-1,測(cè)定菌體干重和培養(yǎng)基中酶活變化,培養(yǎng)基中酶活不再上升時(shí),發(fā)酵過(guò)程結(jié)束。在此條件下進(jìn)行發(fā)酵培養(yǎng),能夠使胞內(nèi)定位目標(biāo)蛋白分泌到胞外。
有益效果:本發(fā)明通過(guò)共表達(dá)磷脂酶C,使大腸桿菌細(xì)胞膜透性提高,可以使胞內(nèi)定位蛋白以非特異滲漏的方式釋放到細(xì)胞外,篩選獲得高性能的磷脂酶C和目標(biāo)蛋白胞外酶活較高的共表達(dá)重組菌。蔗糖磷酸化酶與磷脂酶C共表達(dá)的重組菌胞外蔗糖磷酸化酶的酶活為1445.1U·mL-1,而單獨(dú)表達(dá)的重組菌胞外未檢測(cè)到蔗糖磷酸化酶酶活;海藻糖酶與磷脂酶C共表達(dá)的重組菌胞外海藻糖酶的酶活為322.3U·mL-1,而單獨(dú)表達(dá)的重組菌胞外未檢測(cè)到海藻糖酶酶活。在食品、化妝品及醫(yī)藥行業(yè)有廣泛的應(yīng)用價(jià)值,國(guó)內(nèi)外市場(chǎng)需求較大。
附圖說(shuō)明
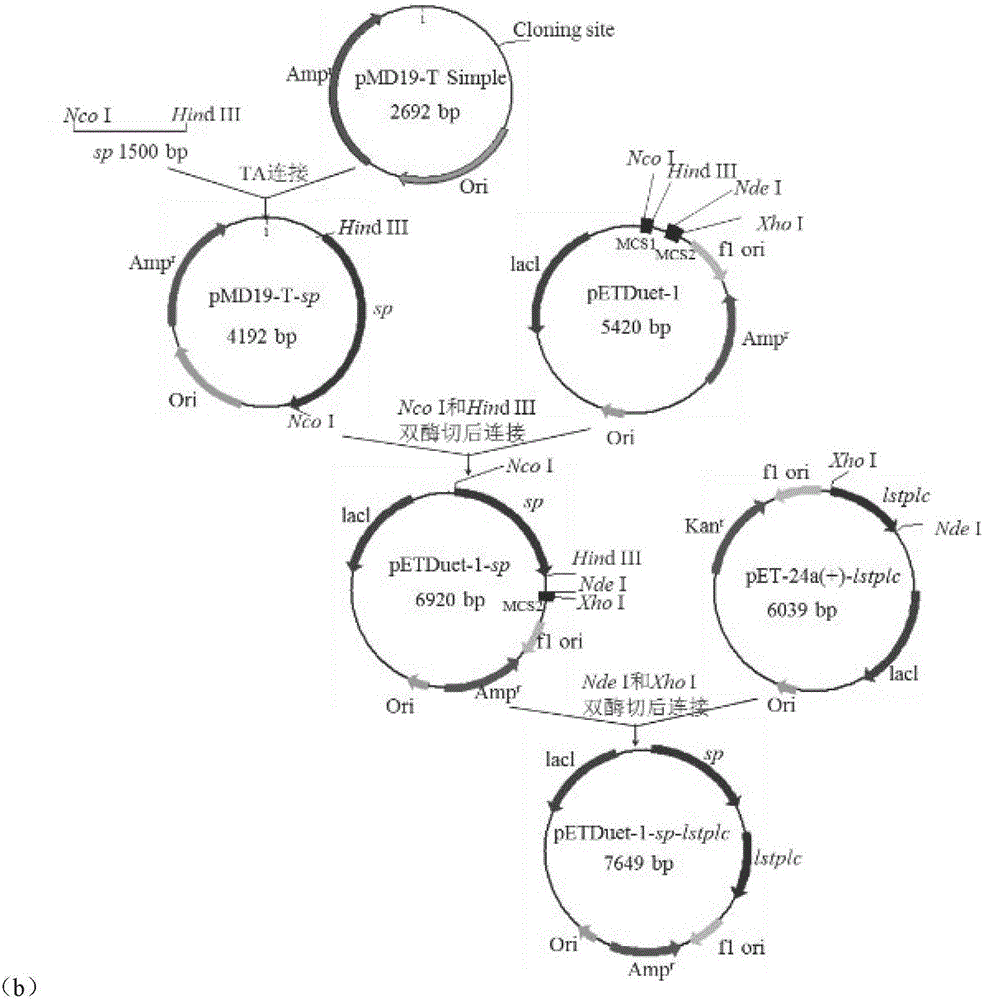
圖1為重組質(zhì)粒的構(gòu)建過(guò)程;a,pETDuet-1-lstplc-sp;b,pETDuet-1-sp-lstplc;
圖2為重組菌的生物量和產(chǎn)酶曲線;其中,a,E.coli BL21(DE3)/pETDuet-1-lstplc-sp;b,E.coli BL21(DE3)/pETDuet-1-sp-lstplc;
圖3為重組菌SDS-PAGE凝膠電泳圖;其中,a,E.coli BL21(DE3)/pETDuet-1-lstplc-sp;b,E.coli BL21(DE3)/pETDuet-1-sp-lstplc;M,Marker;1-3分別為E.coli BL21(DE3)/pETDuet-1-lstplc-sp的發(fā)酵上清液、破壁上清液和破壁沉淀;4-6分別為E.coli BL21(DE3)/pETDuet-1-sp-lstplc的發(fā)酵上清液、破壁上清液和破壁沉淀;
圖4為重組菌的生物量和產(chǎn)酶曲線;其中,a,E.coli BL21(DE3)/pETDuet-1-cpplc-sp;b,E.coli BL21(DE3)/pETDuet-1-sp-cpplc;■,菌體干重;▲,胞外上清酶活;●,總酶活;
圖5為重組菌的SDS-PAGE凝膠電泳圖;其中,a,E.coli BL21(DE3)/pETDuet-1-cpplc-sp;b,E.coli BL21(DE3)/pETDuet-1-sp-cpplc;M,Marker;1-3分別為重組菌E.coli BL21(DE3)/pETDuet-1-cpplc-sp的發(fā)酵上清液,破壁上清液和破壁沉淀;4-6分別為重組菌E.coli BL21(DE3)/pETDuet-1-sp-cpplc的發(fā)酵上清液,破壁上清液和破壁沉淀
圖6為重組菌的生物量和產(chǎn)酶曲線;其中,a,E.coli BL21(DE3)/pETDuet-1-bcplc-sp;b,E.coli BL21(DE3)/pETDuet-1-sp-bcplc;■,菌體干重;▲,胞外上清酶活;●,總酶活;
圖7為重組菌的SDS-PAGE凝膠電泳圖;其中,a,E.coli BL21(DE3)/pETDuet-1-bcplc-sp;b,E.coli BL21(DE3)/pETDuet-1-sp-bcplc;M,Marker;1-3分別為重組菌E.coli BL21(DE3)/pETDuet-1-bcplc-sp的發(fā)酵上清液,破壁上清液和破壁沉淀;4-6分別為重組菌E.coli BL21(DE3)/pETDuet-1-sp-bcplc的發(fā)酵上清液,破壁上清液和破壁沉淀。
圖8為重組菌E.coli BL21(DE3)/pETDuet-1-sp-bcplc高密度發(fā)酵的生物量和產(chǎn)酶曲線;其中,■,菌體干重;▲,胞外上清酶活;
圖9為重組菌E.coli BL21(DE3)/pETDuet-1-sp-bcplc高密度發(fā)酵上清SDS-PAGE凝膠電泳分析(將發(fā)酵上清液稀釋10倍進(jìn)行上樣);其中M,Marker;1-9分別為重組菌E.coli BL21(DE3)/pETDuet-1-sp-bcplc誘導(dǎo)15h、18h、21h、24h、27h、30h、33h、36h、39h發(fā)酵上清液;
圖10為重組菌E.coli BL21(DE3)/pETDuet-1-Tre F-bcplc的生物量和產(chǎn)酶曲線;其中,■,菌體干重;▲,胞外上清酶活;●,總酶活;
圖11為重組菌E.coli BL21(DE3)/pETDuet-1-Tre F-bcplc的SDS-PAGE凝膠電泳分析;其中M,Marker;1-31-3分別為重組菌的發(fā)酵上清液,破壁上清液和破壁沉淀;
圖12為重組菌E.coli BL21(DE3)/pETDuet-1-Tre F-bcplc高密度發(fā)酵的生物量和產(chǎn)酶曲線;其中,■,菌體干重;▲,胞外上清酶活;
具體實(shí)施方式
培養(yǎng)基見(jiàn)表1:
表1培養(yǎng)基成分及配制方法
在具體實(shí)施方式中,以蔗糖磷酸化酶和海藻糖酶為例實(shí)施本發(fā)明的技術(shù)方案,其中蔗糖磷酸化酶的氨基酸序列如SEQ ID NO.7所示,海藻糖酶的氨基酸序列如SEQ ID NO.8所示。
實(shí)施例1:L.monocytogenes磷脂酶C和蔗糖磷酸化酶共表達(dá)
1.重組菌的構(gòu)建:分別以含有L.monocytogenes磷脂酶C基因(氨基酸序列如SEQ ID NO.1所示)的表達(dá)質(zhì)粒pET-24a(+)-lstplc和實(shí)驗(yàn)室保藏的含有蔗糖磷酸化酶基因的表達(dá)質(zhì)粒pET-24a(+)-sp為模板,PCR擴(kuò)增得到含有Nco I和Hind III酶切位點(diǎn)的磷脂酶C基因lstplc和蔗糖磷酸化酶基因sp,然后按照?qǐng)D1(a)和(b)質(zhì)粒構(gòu)建過(guò)程將L.monocytogenes磷脂酶C和蔗糖磷酸化酶基因分別插入到pETDuet-1(購(gòu)自Novagen公司)表達(dá)載體的多克隆位點(diǎn)MCS1或MCS2中,連接產(chǎn)物轉(zhuǎn)化克隆宿主E.coli JM109涂布LB-Amp固體培養(yǎng)基,37℃培養(yǎng)箱培養(yǎng)8h,挑取單菌落,經(jīng)LB-Amp液體培養(yǎng)基37℃培養(yǎng)8h后,收集菌體抽提質(zhì)粒,酶切驗(yàn)證,然后對(duì)驗(yàn)證正確的重組質(zhì)粒測(cè)定DNA序列,獲得重組表達(dá)質(zhì)粒pETDuet-1-lstplc-sp和pETDuet-1-sp-lstplc。將重組表達(dá)質(zhì)粒分別轉(zhuǎn)化E.coli BL21(DE3)感受態(tài)細(xì)胞,涂布LB-Amp固體培養(yǎng)基,37℃培養(yǎng)箱培養(yǎng)12h,挑取單菌落,經(jīng)LB-Amp液體培養(yǎng)基37℃培養(yǎng)8h后保甘油菌,即得到重組菌E.coli BL21(DE3)/pETDuet-1-lstplc-sp和E.coli BL21(DE3)/pETDuet-1-sp-lstplc。
2.重組菌的搖瓶發(fā)酵:將重組菌E.coli BL21(DE3)/pETDuet-1-lstplc-sp和E.coli BL21(DE3)/pETDuet-1-sp-lstplc用移液槍吸取10μL菌液分別接種于10mL LB-Amp液體培養(yǎng)基中,置于37℃恒溫?fù)u床,200r·min-1,培養(yǎng)8-10h,作為種子液。
將培養(yǎng)好的種子液以5%的接種量接入50mL TB-Amp培養(yǎng)基中,置于37℃恒溫?fù)u床,轉(zhuǎn)速200r·min-1,培養(yǎng)2h;加入終濃度為0.4mmol·L-1的IPTG(異丙基硫代-β-D半乳糖苷)進(jìn)行誘導(dǎo)發(fā)酵,25℃,轉(zhuǎn)速200r·min-1,培養(yǎng)20-30h。如圖2(a),重組菌E.coli BL21(DE3)/pETDuet-1-lstplc-sp經(jīng)IPTG誘導(dǎo)發(fā)酵21h后,發(fā)酵上清中蔗糖磷酸化酶的酶活達(dá)到最高為72U·mL-1,占胞內(nèi)總酶活的92.1%。如圖2(b),重組菌E.coli BL21(DE3)/pETDuet-1-sp-lstplc經(jīng)IPTG誘導(dǎo)發(fā)酵26h后,胞外上清的蔗糖磷酸化酶酶活達(dá)到最高為122.2U·mL-1,占總酶活的90.5%。此時(shí),兩株重組菌的磷脂酶C的酶活分別為9.5U·mL-1和9.1U·mL-1。
實(shí)施例2:C.perfringens磷脂酶C和蔗糖磷酸化酶共表達(dá)
1.重組菌的構(gòu)建:分別以含有C.perfringens磷脂酶C基因(氨基酸序列如SEQ ID NO.2所示)的表達(dá)質(zhì)粒pET-24a(+)-cpplc和實(shí)驗(yàn)室保藏的含有蔗糖磷酸化酶基因的表達(dá)質(zhì)粒pET-24a(+)-sp為模板,PCR擴(kuò)增得到含有Nco I和Hind III酶切位點(diǎn)的磷脂酶C基因cpplc和蔗糖磷酸化酶基因sp,然后參照?qǐng)D1(a)和(b)質(zhì)粒構(gòu)建過(guò)程將C.perfringens磷脂酶C和蔗糖磷酸化酶基因分別插入到pETDuet-1表達(dá)載體的多克隆位點(diǎn)MCS1或MCS2中,連接產(chǎn)物轉(zhuǎn)化克隆宿主E.coli JM109涂布LB-Amp固體培養(yǎng)基,37℃培養(yǎng)箱培養(yǎng)8h,挑取單菌落,經(jīng)LB-Amp液體培養(yǎng)基37℃培養(yǎng)8h后,收集菌體抽提質(zhì)粒,酶切驗(yàn)證,然后對(duì)驗(yàn)證正確的重組質(zhì)粒測(cè)定DNA序列,獲得重組表達(dá)質(zhì)粒pETDuet-1-cpplc-sp和pETDuet-1-sp-cpplc。將重組表達(dá)質(zhì)粒分別轉(zhuǎn)化E.coli BL21(DE3)感受態(tài)細(xì)胞,涂布LB-Amp固體培養(yǎng)基,37℃培養(yǎng)箱培養(yǎng)12h,挑取單菌落,經(jīng)LB-Amp液體培養(yǎng)基37℃培養(yǎng)8h后保甘油菌,即得到重組菌E.coli BL21(DE3)/pETDuet-1-cpplc-sp和E.coli BL21(DE3)/pETDuet-1-sp-cpplc。
2.重組菌的搖瓶發(fā)酵:培養(yǎng)方法同實(shí)施例1。如圖4(a),重組菌E.coli BL21(DE3)/pETDuet-1-cpplc-sp誘導(dǎo)發(fā)酵26h時(shí),發(fā)酵液上清中蔗糖磷酸化酶的酶活達(dá)到最高為16.2U·mL-1,總酶活為187.2U·mL-1,胞外酶活占總酶活的8.6%。如圖4(b),重組菌E.coli BL21(DE3)/pETDuet-1-sp-cpplc誘導(dǎo)發(fā)酵30h時(shí),胞外上清中蔗糖磷酸化酶的酶活達(dá)到最高為19.3U·mL-1,總酶活為285.0U·mL-1,胞外酶活約占總酶活的6.7%。兩種重組菌磷脂酶C的總酶活分別為8.4U·mL-1和8.0U·mL-1。
實(shí)施例3:B.cereus磷脂酶C和蔗糖磷酸化酶共表達(dá)
1.重組菌的構(gòu)建:分別以含有B.cereus磷脂酶C基因(氨基酸序列如SEQ ID NO.3所示)的表達(dá)質(zhì)粒pET-24a(+)-bcplc和實(shí)驗(yàn)室保藏的含有蔗糖磷酸化酶基因的表達(dá)質(zhì)粒pET-24a(+)-sp為模板,PCR擴(kuò)增得到含有Nco I和Hind III酶切位點(diǎn)的磷脂酶C基因bcplc和蔗糖磷酸化酶基因sp,然后參照?qǐng)D1(a)和(b)質(zhì)粒構(gòu)建過(guò)程將B.cereus磷脂酶C和蔗糖磷酸化酶基因分別插入到pETDuet-1表達(dá)載體的多克隆位點(diǎn)MCS1或MCS2中,連接產(chǎn)物轉(zhuǎn)化克隆宿主E.coli JM109涂布LB-Amp固體培養(yǎng)基,37℃培養(yǎng)箱培養(yǎng)8h,挑取單菌落,經(jīng)LB-Amp液體培養(yǎng)基37℃培養(yǎng)8h后,收集菌體抽提質(zhì)粒,酶切驗(yàn)證,然后對(duì)驗(yàn)證正確的重組質(zhì)粒測(cè)定DNA序列,獲得重組表達(dá)質(zhì)粒pETDuet-1-bcplc-sp和pETDuet-1-sp-bcplc。將重組表達(dá)質(zhì)粒分別轉(zhuǎn)化E.coli BL21(DE3)感受態(tài)細(xì)胞,涂布LB-Amp固體培養(yǎng)基,37℃培養(yǎng)箱培養(yǎng)12h,挑取單菌落,經(jīng)LB-Amp液體培養(yǎng)基37℃培養(yǎng)8h后保甘油菌,即得到重組菌E.coli BL21(DE3)/pETDuet-1-bcplc-sp和E.coli BL21(DE3)/pETDuet-1-sp-bcplc。
2.重組菌的搖瓶發(fā)酵:培養(yǎng)方法同實(shí)施例1。如圖6(a),重組菌E.coli BL21(DE3)/pETDuet-1-bcplc-sp在誘導(dǎo)發(fā)酵28h時(shí),發(fā)酵上清中的蔗糖磷酸化酶酶活達(dá)到最高為84.2U·mL-1,此時(shí)總酶活為101.5U·mL-1,胞外酶活約占總酶活的82.9%;如圖6(b),重組菌E.coli BL21(DE3)/pETDuet-1-sp-bcplc在誘導(dǎo)發(fā)酵28h時(shí),胞外上清中蔗糖磷酸化酶酶活達(dá)到最高為174U·mL-1,總酶活為224.5U·mL-1,胞外酶活約占總酶活的77.5%。兩種重組菌磷脂酶C的總酶活分別為18.5U·mL-1和13.3U·mL-1。
不同的磷脂酶C和蔗糖磷酸化酶共表達(dá)重組菌的搖瓶發(fā)酵生長(zhǎng)和產(chǎn)酶情況如表2所示,當(dāng)L.monocytogenes磷脂酶C和蔗糖磷酸化酶共表達(dá)時(shí),重組菌蔗糖磷酸化酶的胞外產(chǎn)酶比例最高,同L.monocytogenes磷脂酶C單獨(dú)表達(dá)結(jié)果類似,但是磷脂酶C對(duì)蔗糖磷酸化酶的表達(dá)具有很大的影響,與另外兩種磷脂酶C和蔗糖磷酸化酶共表達(dá)相比,蔗糖磷酸化酶的發(fā)酵周期相對(duì)較短,總酶活最低;當(dāng)C.perfringens磷脂酶C和蔗糖磷酸化酶共表達(dá)時(shí),磷脂酶C對(duì)蔗糖磷酸化酶的表達(dá)影響較小,重組菌發(fā)酵周期較長(zhǎng),蔗糖磷酸化酶的總酶活最高,但是由于C.perfringens磷脂酶C對(duì)細(xì)胞膜磷脂成分磷脂酰乙醇胺的水解活性較差,導(dǎo)致蔗糖磷酸化酶的釋放到胞外的比例很低;與另外兩種磷脂酶C和蔗糖磷酸化酶共表達(dá)相比,當(dāng)B.cereus磷脂酶C與蔗糖磷酸化酶共表達(dá)時(shí),蔗糖磷酸化酶的胞外酶活最高,重組菌E.coli BL21(DE3)/pETDuet-1-sp-bcplc蔗糖磷酸化酶的胞外酶活為174.0U·mL-1,由于磷脂酶C對(duì)細(xì)胞膜磷脂成分的水解作用,蔗糖磷酸化酶的胞外產(chǎn)酶比例也可到達(dá)77.5%。
表2
注:lstplc-sp、sp-lstplc、cpplc-sp、sp-cpplc、bcplc-sp和sp-bcplc分別表示重組菌E.coli BL21(DE3)/pETDuet-1-lstplc-sp、E.coli BL21(DE3)/pETDuet-1-sp-lstplc、E.coli BL21(DE3)/pETDuet-1-cpplc-sp、E.coli BL21(DE3)/pETDuet-1-sp-cpplc、E.coli BL21(DE3)/pETDuet-1-bcplc-sp和E.coli BL21(DE3)/pETDuet-1-sp-bcplc。
綜上,根據(jù)不同磷脂酶C和蔗糖磷酸化酶共表達(dá)對(duì)蔗糖磷酸化酶的酶活和胞外產(chǎn)酶比例的比較分析,重組菌E.coli BL21(DE3)/pETDuet-1-sp-bcplc蔗糖磷酸化酶的胞外酶活最高,具有更好的工業(yè)應(yīng)用價(jià)值。
實(shí)施例4:重組菌E.coli BL21(DE3)/pETDuet-1-sp-bcplc的3L發(fā)酵罐高密度發(fā)酵
種子培養(yǎng):將-80℃甘油管保存的E.coli BL21(DE3)/pETDuet-1-sp-bcplc菌株接入種子培養(yǎng)基中,使用恒溫?fù)u床進(jìn)行培養(yǎng),溫度37℃,轉(zhuǎn)速200rpm,培養(yǎng)8h。
發(fā)酵產(chǎn)酶:將種子液以8%接種量接入發(fā)酵培養(yǎng)基。通過(guò)控制攪拌轉(zhuǎn)速和通風(fēng)量使溶氧維持30%,控制溫度為37℃,流加氨水控制pH在7.0。待最初的甘油消耗完后,以比生長(zhǎng)速率μ=0.2h-1指數(shù)流加的方式補(bǔ)加補(bǔ)料培養(yǎng)基。當(dāng)細(xì)胞濃度達(dá)到OD600=30時(shí),開(kāi)始以0.2g·L-1·h-1流速恒速流加乳糖進(jìn)行誘導(dǎo),誘導(dǎo)溫度30℃,誘導(dǎo)36h左右。發(fā)酵培養(yǎng)結(jié)束后,30℃誘導(dǎo)39h,蔗糖磷酸化酶的酶活力最高可以達(dá)到1445.1U·mL-1,占總酶活的81.6%
實(shí)施例5:蔗糖磷酸化酶在E.coli BL21(DE3)單獨(dú)表達(dá)
本發(fā)明將蔗糖磷酸化酶基因連接在pET-24a(+)表達(dá)載體上,構(gòu)建了蔗糖磷酸化酶的單獨(dú)表達(dá)重組菌E.coli BL21(DE3)/pET-24a(+)-sp,對(duì)重組菌進(jìn)行搖瓶發(fā)酵,在發(fā)酵上清中未檢測(cè)到蔗糖磷酸化酶的酶活。
實(shí)施例6:海藻糖酶與B.cereus磷脂酶C共表達(dá)
1.重組菌的構(gòu)建:分別以含有B.cereus磷脂酶C基因(氨基酸序列如SEQ ID NO.3所示)的表達(dá)質(zhì)粒pET-24a(+)-bcplc和實(shí)驗(yàn)室保藏的含有海藻糖酶基因的表達(dá)質(zhì)粒pET-24a(+)-Tre F(氨基酸序列如SEQ ID NO.8所示)為模板,參照?qǐng)D1(b)的質(zhì)粒構(gòu)建過(guò)程將將海藻糖酶基因Tre F和B.cereus磷脂酶C基因bcplc分別連接到pETDuet-1表達(dá)載體的多克隆位點(diǎn)MCS1和MCS2,連接產(chǎn)物轉(zhuǎn)化克隆宿主E.coli JM109涂布LB-Amp固體培養(yǎng)基,37℃培養(yǎng)箱培養(yǎng)8h,挑取單菌落,經(jīng)LB-Amp液體培養(yǎng)基37℃培養(yǎng)8h后,收集菌體抽提質(zhì)粒,酶切驗(yàn)證,然后對(duì)驗(yàn)證正確的重組質(zhì)粒測(cè)定DNA序列,獲得重組表達(dá)質(zhì)粒pETDuet-1-Tre F-bcplc。將重組表達(dá)質(zhì)粒分別轉(zhuǎn)化E.coli BL21(DE3)感受態(tài)細(xì)胞,涂布LB-Amp固體培養(yǎng)基,37℃培養(yǎng)箱培養(yǎng)12h,挑取單菌落,經(jīng)LB-Amp液體培養(yǎng)基37℃培養(yǎng)8h后保甘油菌,即得到重組菌E.coli BL21(DE3)/pETDuet-1-Tre F-bcplc。
2.重組菌的搖瓶發(fā)酵:培養(yǎng)方法同實(shí)施例1。如圖11,重組菌E.coli BL21(DE3)/pETDuet-1-Tre F-bcplc重組菌胞外酶活達(dá)到最高為64.6U·mL-1,總酶活為85.9U·mL-1,胞外酶活約占總酶活的75.2%;此時(shí),磷脂酶C胞內(nèi)外酶活為12.5U·mL-1,表明B.cereus磷脂酶C共表達(dá)海藻糖酶時(shí)可以使海藻糖酶很好的分泌到胞外。
3.重組菌的高密度發(fā)酵:發(fā)酵條件同實(shí)施例4.如圖12,重組菌E.coli BL21(DE3)/pETDuet-1-Tre F-bcplc發(fā)酵上清中海藻糖酶活最高為322.3U·mL-1,胞外酶活約占總酶活的40.6%。
實(shí)施例7:海藻糖酶在E.coli BL21(DE3)單獨(dú)表達(dá)
本發(fā)明將海藻糖酶基因與pET-24a(+)表達(dá)載體連接,構(gòu)建了蔗糖磷酸化酶的單獨(dú)表達(dá)重組菌E.coli BL21(DE3)/pET-24a(+)-Tre F,對(duì)重組菌進(jìn)行搖瓶發(fā)酵,在發(fā)酵上清中未檢測(cè)到海藻糖酶的酶活。
雖然本發(fā)明已以較佳實(shí)施例公開(kāi)如上,但其并非用以限定本發(fā)明,任何熟悉此技術(shù)的人,在不脫離本發(fā)明的精神和范圍內(nèi),都可做各種的改動(dòng)與修飾,因此本發(fā)明的保護(hù)范圍應(yīng)該以權(quán)利要求書所界定的為準(zhǔn)。
SEQUENCE LISTING
<110> 江南大學(xué)
<120> 一種應(yīng)用磷脂酶C將胞內(nèi)蛋白在胞外表達(dá)的方法
<160> 8
<170> PatentIn version 3.3
<210> 1
<211> 239
<212> PRT
<213> 人工序列
<400> 1
Met Trp Ser Ala Asp Asn Pro Thr Asn Thr Asp Val Asn Thr His Tyr
1 5 10 15
Trp Leu Phe Lys Gln Ala Glu Lys Ile Leu Ala Lys Asp Val Asn His
20 25 30
Met Arg Ala Asn Leu Met Asn Glu Leu Lys Lys Phe Asp Lys Gln Ile
35 40 45
Ala Gln Gly Ile Tyr Asp Ala Asp His Lys Asn Pro Tyr Tyr Asp Thr
50 55 60
Ser Thr Phe Leu Ser His Phe Tyr Asn Pro Asp Arg Asp Asn Thr Tyr
65 70 75 80
Leu Pro Gly Phe Ala Asn Ala Lys Ile Thr Gly Ala Lys Tyr Phe Asn
85 90 95
Gln Ser Val Thr Asp Tyr Arg Glu Gly Lys Phe Asp Thr Ala Phe Tyr
100 105 110
Lys Leu Gly Leu Ala Ile His Tyr Tyr Thr Asp Ile Ser Gln Pro Met
115 120 125
His Ala Asn Asn Phe Thr Ala Ile Ser Tyr Pro Pro Gly Tyr His Cys
130 135 140
Ala Tyr Glu Asn Tyr Val Asp Thr Ile Lys His Asn Tyr Gln Ala Thr
145 150 155 160
Glu Asp Met Val Ala Lys Arg Phe Cys Ser Asp Asp Val Lys Asp Trp
165 170 175
Leu Tyr Glu Asn Ala Lys Arg Ala Lys Ala Asp Tyr Pro Lys Ile Val
180 185 190
Asn Ala Lys Thr Lys Lys Ser Tyr Leu Val Gly Asn Ser Glu Trp Lys
195 200 205
Lys Asp Thr Val Glu Pro Thr Gly Ala Arg Leu Arg Asp Ser Gln Gln
210 215 220
Thr Leu Ala Gly Phe Leu Glu Phe Trp Ser Lys Lys Thr Asn Glu
225 230 235
<210> 2
<211> 371
<212> PRT
<213> 人工序列
<400> 2
Met Trp Asp Gly Lys Ile Asp Gly Thr Gly Thr His Ala Met Ile Val
1 5 10 15
Thr Gln Gly Val Ser Ile Leu Glu Asn Asp Leu Ser Lys Asn Glu Pro
20 25 30
Glu Ser Val Arg Lys Asn Leu Glu Ile Leu Lys Glu Asn Met His Glu
35 40 45
Leu Gln Leu Gly Ser Thr Tyr Pro Asp Tyr Asp Lys Asn Ala Tyr Asp
50 55 60
Leu Tyr Gln Asp His Phe Trp Asp Pro Asp Thr Asp Asn Asn Phe Ser
65 70 75 80
Lys Asp Asn Ser Trp Tyr Leu Ala Tyr Ser Ile Pro Asp Thr Gly Glu
85 90 95
Ser Gln Ile Arg Lys Phe Ser Ala Leu Ala Arg Tyr Glu Trp Gln Arg
100 105 110
Gly Asn Tyr Lys Gln Ala Thr Phe Tyr Leu Gly Glu Ala Met His Tyr
115 120 125
Phe Gly Asp Ile Asp Thr Pro Tyr His Pro Ala Asn Val Thr Ala Val
130 135 140
Asp Ser Ala Gly His Val Lys Phe Glu Thr Phe Ala Glu Glu Arg Lys
145 150 155 160
Glu Gln Tyr Lys Ile Asn Thr Ala Gly Cys Lys Thr Asn Glu Ala Phe
165 170 175
Tyr Thr Asp Ile Leu Lys Asn Lys Asp Phe Asn Ala Trp Ser Lys Glu
180 185 190
Tyr Ala Arg Gly Phe Ala Lys Thr Gly Lys Ser Ile Tyr Tyr Ser His
195 200 205
Ala Ser Met Ser His Ser Trp Asp Asp Trp Asp Tyr Ala Ala Lys Val
210 215 220
Thr Leu Ala Asn Ser Gln Lys Gly Thr Ala Gly Tyr Ile Tyr Arg Phe
225 230 235 240
Leu His Asp Val Ser Glu Gly Asn Asp Pro Ser Val Gly Lys Asn Val
245 250 255
Lys Glu Leu Val Ala Tyr Ile Ser Thr Ser Gly Glu Lys Asp Ala Gly
260 265 270
Thr Asp Asp Tyr Met Tyr Phe Gly Ile Lys Thr Lys Asp Gly Lys Thr
275 280 285
Gln Glu Trp Glu Met Asp Asn Pro Gly Asn Asp Phe Met Thr Gly Ser
290 295 300
Lys Asp Thr Tyr Thr Phe Lys Leu Lys Asp Glu Asn Leu Lys Ile Asp
305 310 315 320
Asp Ile Gln Asn Met Trp Ile Arg Lys Arg Lys Tyr Thr Ala Phe Ser
325 330 335
Asp Ala Tyr Lys Pro Glu Asn Ile Lys Ile Ile Ala Asn Gly Lys Val
340 345 350
Val Val Asp Lys Asp Ile Asn Glu Trp Ile Ser Gly Asn Ser Thr Tyr
355 360 365
Asn Ile Lys
370
<210> 3
<211> 259
<212> PRT
<213> 人工序列
<400> 3
His Glu Asn Asp Gly Gly Ser Lys Ile Lys Ile Val His Arg Trp Ser
1 5 10 15
Ala Glu Asp Lys His Lys Glu Gly Val Asn Ser His Leu Trp Ile Val
20 25 30
Asn Arg Ala Ile Asp Ile Met Ser Arg Asn Thr Thr Leu Val Lys Gln
35 40 45
Asp Arg Val Ala Gln Leu Asn Glu Trp Arg Thr Glu Leu Glu Asn Gly
50 55 60
Ile Tyr Ala Ala Asp Tyr Glu Asn Pro Tyr Tyr Asp Asn Ser Thr Phe
65 70 75 80
Ala Ser His Phe Tyr Asp Pro Asp Asn Gly Lys Thr Tyr Ile Pro Phe
85 90 95
Ala Lys Gln Ala Lys Glu Thr Gly Ala Lys Tyr Phe Lys Leu Ala Gly
100 105 110
Glu Ser Tyr Lys Asn Lys Asp Met Lys Gln Ala Phe Phe Tyr Leu Gly
115 120 125
Leu Ser Leu His Tyr Leu Gly Asp Val Asn Gln Pro Met His Ala Ala
130 135 140
Asn Phe Thr Asn Leu Ser Tyr Pro Gln Gly Phe His Ser Lys Tyr Glu
145 150 155 160
Asn Phe Val Asp Thr Ile Lys Asp Asn Tyr Lys Val Thr Asp Gly Asn
165 170 175
Gly Tyr Trp Asn Trp Lys Gly Thr Asn Pro Glu Glu Trp Ile His Gly
180 185 190
Ala Ala Val Val Ala Lys Gln Asp Tyr Ser Gly Ile Val Asn Asp Asn
195 200 205
Thr Lys Asp Trp Phe Val Lys Ala Ala Val Ser Gln Glu Tyr Ala Asp
210 215 220
Lys Trp Arg Ala Glu Val Thr Pro Met Thr Gly Lys Arg Leu Met Asp
225 230 235 240
Ala Gln Arg Val Thr Ala Gly Tyr Ile Gln Leu Trp Phe Asp Thr Tyr
245 250 255
Gly Asp Arg
<210> 4
<211> 720
<212> DNA
<213> 人工序列
<400> 4
atgtggtcag cagataatcc gaccaatacc gatgtgaata cccattattg gctcttcaag 60
caggcagaaa aaattctggc aaaagatgtt aatcacatgc gcgcaaatct gatgaatgaa 120
ctgaaaaaat ttgataaaca gattgcacag ggtatttatg atgcagatca taaaaatccg 180
tattatgata cctcaacctt tctgtctcat ttttataatc cagatcgcga taatacctat 240
ctgcctggtt ttgcaaatgc aaaaattacc ggcgcaaaat attttaatca gagtgtgacc 300
gattatcgcg aaggtaaatt tgataccgca ttttataaac tgggcctggc aattcattat 360
tataccgata ttagtcagcc gatgcacgca aataatttta ccgcaattag ctatccgcca 420
ggttatcatt gtgcttatga aaattatgtg gataccatta aacataatta tcaggcaacc 480
gaagatatgg tggcaaaacg cttttgtagc gatgatgtta aagattggct gtatgaaaat 540
gcaaaacgtg caaaagcaga ttatccgaaa attgttaatg caaaaaccaa aaaatcttat 600
ctggtgggta atagcgaatg gaaaaaagat accgtggaac cgaccggcgc acgcctgcgt 660
gattcacagc agaccctggc aggctttctg gaattttgga gtaaaaaaac caatgaataa 720
<210> 5
<211> 1116
<212> DNA
<213> 人工序列
<400> 5
atgtgggatg gtaaaattga tgggaccggc acccatgcaa tgattgtgac ccagggtgtg 60
tcaattctgg aaaatgatct gtctaaaaat gaaccggaat cagttcgtaa aaatctggaa 120
attctgaaag aaaatatgca cgaactgcag ctgggctcta cctaccctga ttatgataaa 180
aatgcctacg atctgtatca ggatcatttt tgggaccccg ataccgataa taatttttct 240
aaagataata gttggtatct ggcatatagc atccccgata ccggcgaatc tcagattcgc 300
aaattttcag cactggcacg ctatgaatgg cagcgcggta attataaaca ggcaaccttt 360
tatctgggcg aagcaatgca ctattttggc gatattgata ccccgtatca tccggcaaat 420
gtgaccgcag ttgatagcgc aggccatgtt aaatttgaaa cctttgcaga agaacgcaaa 480
gaacagtata aaatcaacac cgcaggttgt aaaaccaatg aagcatttta taccgatatt 540
ctgaaaaata aagattttaa tgcttggtct aaagaatatg cacgcggctt tgcaaaaacc 600
ggtaaatcaa tttattatag tcatgcaagt atgtctcata gttgggatga ttgggattat 660
gcagcaaaag tgaccctggc aaatagccag aaaggtacag caggctatat ttatcgcttt 720
ctgcatgatg ttagcgaagg taatgatccg agcgtgggca aaaatgttaa agaactggtt 780
gcatatatta gtaccagtgg cgaaaaagat gcaggcaccg atgattatat gtattttggt 840
attaaaacca aagatggcaa aacccaggaa tgggaaatgg ataatcctgg aaatgatttt 900
atgaccggct ctaaagatac ctataccttt aaactgaaag atgaaaatct gaaaattgat 960
gatattcaga atatgtggat tcgcaaacgt aaatataccg catttagcga tgcgtataaa 1020
ccggaaaata ttaaaattat tgcaaatggc aaagtggttg tggataaaga tattaacgaa 1080
tggattagcg gtaattcaac ctataatatt aaataa 1116
<210> 6
<211> 780
<212> DNA
<213> 人工序列
<400> 6
catgaaaatg atgggggaag taaaataaaa atagttcacc gctggtctgc tgaagataaa 60
cataaagaag gtgtaaattc tcatttatgg attgtaaacc gtgcgattga tattatgtct 120
cgcaatacaa cacttgtaaa acaagatcga gttgcacaat taaatgaatg gcgtacggag 180
ttagagaacg gtatttatgc tgctgactat gaaaatcctt attatgataa tagcacattt 240
gcttcacatt tctatgatcc agacaatgga aaaacatata ttccatttgc aaagcaggca 300
aaagaaactg gcgctaaata ttttaaatta gctggtgaat catacaaaaa taaagatatg 360
aaacaagcat tcttctattt aggattatct cttcattatt taggagatgt aaaccaaccg 420
atgcatgcgg caaactttac aaatctttca tatccacaag gattccattc taaatatgaa 480
aactttgtag atacgataaa agataattat aaagtaacgg atggaaatgg atattggaac 540
tggaaaggta caaatccaga agagtggatt catggagcgg cagtagtagc gaaacaagat 600
tactctggaa ttgtaaatga taatacgaaa gattggttcg taaaagcagc tgtgtcacaa 660
gaatatgcag ataaatggcg cgctgaagtt acaccaatga caggtaagcg attaatggat 720
gcacaacgtg ttactgctgg atacattcag ctttggtttg atacgtacgg agatcgttaa 780
<210> 7
<211> 490
<212> PRT
<213> 人工序列
<400> 7
Met Glu Ile Gln Asn Lys Ala Met Leu Ile Thr Tyr Ala Asp Ser Leu
1 5 10 15
Gly Lys Asn Leu Lys Asp Val His Gln Val Leu Lys Glu Asp Ile Gly
20 25 30
Asp Ala Ile Gly Gly Val His Leu Leu Pro Phe Phe Pro Ser Thr Gly
35 40 45
Asp Arg Gly Phe Ala Pro Ala Asp Tyr Thr Arg Val Asp Ala Ala Phe
50 55 60
Gly Asp Trp Ala Asp Val Glu Ala Leu Gly Glu Glu Tyr Tyr Leu Met
65 70 75 80
Phe Asp Phe Met Ile Asn His Ile Ser Arg Glu Ser Val Met Tyr Gln
85 90 95
Asp Phe Lys Lys Asn His Asp Asp Ser Lys Tyr Lys Asp Phe Phe Ile
100 105 110
Arg Trp Glu Lys Phe Trp Ala Lys Ala Gly Glu Asn Arg Pro Thr Gln
115 120 125
Ala Asp Val Asp Leu Ile Tyr Lys Arg Lys Asp Lys Ala Pro Thr Gln
130 135 140
Glu Ile Thr Phe Asp Asp Gly Thr Thr Glu Asn Leu Trp Asn Thr Phe
145 150 155 160
Gly Glu Glu Gln Ile Asp Ile Asp Val Asn Ser Ala Ile Ala Lys Glu
165 170 175
Phe Ile Lys Thr Thr Leu Glu Asp Met Val Lys His Gly Ala Asn Leu
180 185 190
Ile Arg Leu Asp Ala Phe Ala Tyr Ala Val Lys Lys Val Asp Thr Asn
195 200 205
Asp Phe Phe Val Glu Pro Glu Ile Trp Asp Thr Leu Asn Glu Val Arg
210 215 220
Glu Ile Leu Thr Pro Leu Lys Ala Glu Ile Leu Pro Glu Ile His Glu
225 230 235 240
His Tyr Ser Ile Pro Lys Lys Ile Asn Asp His Gly Tyr Phe Thr Tyr
245 250 255
Asp Phe Ala Leu Pro Met Thr Thr Leu Tyr Thr Leu Tyr Ser Gly Lys
260 265 270
Thr Asn Gln Leu Ala Lys Trp Leu Lys Met Ser Pro Met Lys Gln Phe
275 280 285
Thr Thr Leu Asp Thr His Asp Gly Ile Gly Val Val Asp Ala Arg Asp
290 295 300
Ile Leu Thr Asp Asp Glu Ile Asp Tyr Ala Ser Glu Gln Leu Tyr Lys
305 310 315 320
Val Gly Ala Asn Val Lys Lys Thr Tyr Ser Ser Ala Ser Tyr Asn Asn
325 330 335
Leu Asp Ile Tyr Gln Ile Asn Ser Thr Tyr Tyr Ser Ala Leu Gly Asn
340 345 350
Asp Asp Ala Ala Tyr Leu Leu Ser Arg Val Phe Gln Val Phe Ala Pro
355 360 365
Gly Ile Pro Gln Ile Tyr Tyr Val Gly Leu Leu Ala Gly Glu Asn Asp
370 375 380
Ile Ala Leu Leu Glu Ser Thr Lys Glu Gly Arg Asn Ile Asn Arg His
385 390 395 400
Tyr Tyr Thr Arg Glu Glu Val Lys Ser Glu Val Lys Arg Pro Val Val
405 410 415
Ala Asn Leu Leu Lys Leu Leu Ser Trp Arg Asn Glu Ser Pro Ala Phe
420 425 430
Asp Leu Ala Gly Ser Ile Thr Val Asp Thr Pro Thr Asp Thr Thr Ile
435 440 445
Val Val Thr Arg Gln Asp Glu Asn Gly Gln Asn Lys Ala Val Leu Thr
450 455 460
Ala Asp Ala Ala Asn Lys Thr Phe Glu Ile Val Glu Asn Gly Gln Thr
465 470 475 480
Val Met Ser Ser Asp Asn Leu Thr Gln Asn
485 490
<210> 8
<211> 549
<212> PRT
<213> 人工序列
<400> 8
Met Leu Asn Gln Lys Ile Gln Asn Pro Asn Pro Asp Glu Leu Met Ile
1 5 10 15
Glu Val Asp Leu Cys Tyr Glu Leu Asp Pro Tyr Glu Leu Lys Leu Asp
20 25 30
Glu Met Ile Glu Ala Glu Pro Glu Pro Glu Met Ile Glu Gly Leu Pro
35 40 45
Ala Ser Asp Ala Leu Thr Pro Ala Asp Arg Tyr Leu Glu Leu Phe Glu
50 55 60
His Val Gln Ser Ala Lys Ile Phe Pro Asp Ser Lys Thr Phe Pro Asp
65 70 75 80
Cys Ala Pro Lys Met Asp Pro Leu Asp Ile Leu Ile Arg Tyr Arg Lys
85 90 95
Val Arg Arg His Arg Asp Phe Asp Leu Arg Lys Phe Val Glu Asn His
100 105 110
Phe Trp Leu Pro Glu Val Tyr Ser Ser Glu Tyr Val Ser Asp Pro Gln
115 120 125
Asn Ser Leu Lys Glu His Ile Asp Gln Leu Trp Pro Val Leu Thr Arg
130 135 140
Glu Pro Gln Asp His Ile Pro Trp Ser Ser Leu Leu Ala Leu Pro Gln
145 150 155 160
Ser Tyr Ile Val Pro Gly Gly Arg Phe Ser Glu Thr Tyr Tyr Trp Asp
165 170 175
Ser Tyr Phe Thr Met Leu Gly Leu Ala Glu Ser Gly Arg Glu Asp Leu
180 185 190
Leu Lys Cys Met Ala Asp Asn Phe Ala Trp Met Ile Glu Asn Tyr Gly
195 200 205
His Ile Pro Asn Gly Asn Arg Thr Tyr Tyr Leu Ser Arg Ser Gln Pro
210 215 220
Pro Val Phe Ala Leu Met Val Glu Leu Phe Glu Glu Asp Gly Val Arg
225 230 235 240
Gly Ala Arg Arg Tyr Leu Asp His Leu Lys Met Glu Tyr Ala Phe Trp
245 250 255
Met Asp Gly Ala Glu Ser Leu Ile Pro Asn Gln Ala Tyr Arg His Val
260 265 270
Val Arg Met Pro Asp Gly Ser Leu Leu Asn Arg Tyr Trp Asp Asp Arg
275 280 285
Asp Thr Pro Arg Asp Glu Ser Trp Leu Glu Asp Val Glu Thr Ala Lys
290 295 300
His Ser Gly Arg Pro Pro Asn Glu Val Tyr Arg Asp Leu Arg Ala Gly
305 310 315 320
Ala Ala Ser Gly Trp Asp Tyr Ser Ser Arg Trp Leu Arg Asp Thr Gly
325 330 335
Arg Leu Ala Ser Ile Arg Thr Thr Gln Phe Ile Pro Ile Asp Leu Asn
340 345 350
Ala Phe Leu Phe Lys Leu Glu Ser Ala Ile Ala Asn Ile Ser Ala Leu
355 360 365
Lys Gly Glu Lys Glu Thr Glu Ala Leu Phe Arg Gln Lys Ala Ser Ala
370 375 380
Arg Arg Asp Ala Val Asn Arg Tyr Leu Trp Asp Asp Glu Asn Gly Ile
385 390 395 400
Tyr Arg Asp Tyr Asp Trp Arg Arg Glu Gln Leu Ala Leu Phe Ser Ala
405 410 415
Ala Ala Ile Val Pro Leu Tyr Val Gly Met Ala Asn His Glu Gln Ala
420 425 430
Asp Arg Leu Ala Asn Ala Val Arg Ser Arg Leu Leu Thr Pro Gly Gly
435 440 445
Ile Leu Ala Ser Glu Tyr Glu Thr Gly Glu Gln Trp Asp Lys Pro Asn
450 455 460
Gly Trp Ala Pro Leu Gln Trp Met Ala Ile Gln Gly Phe Lys Met Tyr
465 470 475 480
Gly Asp Asp Leu Leu Gly Asp Glu Ile Ala Arg Ser Trp Leu Lys Thr
485 490 495
Val Asn Gln Phe Tyr Leu Glu Gln His Lys Leu Ile Glu Lys Tyr His
500 505 510
Ile Ala Asp Gly Val Pro Arg Glu Gly Gly Gly Gly Glu Tyr Pro Leu
515 520 525
Gln Asp Gly Phe Gly Trp Thr Asn Gly Val Val Arg Arg Leu Ile Gly
530 535 540
Leu Tyr Gly Glu Pro
545
- 還沒(méi)有人留言評(píng)論。精彩留言會(huì)獲得點(diǎn)贊!
- 人Lp-PLA2生物素-鏈霉親和素?zé)晒饷庖邔游鰴z測(cè)卡及其制備方法
- 一種硒蛋白磷脂保健品及其制備方法
- 一種脂蛋白磷脂酶a2測(cè)定試劑盒及其使用方法
- 一種重組華支睪吸蟲(chóng)分泌型磷脂酶a2蛋白及其體外可溶性表達(dá)與純化制備方法
- 脂蛋白相關(guān)磷脂酶a2單克隆抗體、抗體對(duì)及制備方法、用圖
- 一種與咪唑乙煙酸抗性相關(guān)蛋白bm-1及其編碼基因與應(yīng)用
- 一種磷脂膠原蛋白豆?jié){粉的制作方法
- 油菜抗旱及ABA耐受相關(guān)的蛋白BnSIP1的基因及其應(yīng)用
- 玉米分泌型磷脂酶A2基因ZmsPLA2-2在改變植物抗旱性中的應(yīng)用
- 脂蛋白磷脂酶A2(Lp-PLA2)免疫熒光探針檢測(cè)試劑盒的制作方法
- 自然酶活性脂蛋白相關(guān)磷脂酶a2及制備過(guò)程的制作方法
- 脂蛋白磷脂酶a2免疫層析定量檢測(cè)試紙條的制作方法
- 一種檢測(cè)脂蛋白磷脂酶a2濃度的試劑盒及其制備方法
- 含脂蛋白相關(guān)性磷脂酶a2抑制劑的藥物組合物及應(yīng)用
- 一種定量檢測(cè)樣本中脂蛋白磷脂酶a2濃度的液態(tài)芯片試劑盒及其制備方法
- 用于檢測(cè)脂蛋白相關(guān)磷脂酶a2的試劑盒及其制備和應(yīng)用
- 同時(shí)測(cè)定血漿中脂蛋白磷脂酶a2和c反應(yīng)蛋白的層析法快速檢測(cè)卡及試劑盒的制作方法
- 血漿脂蛋白磷脂酶a2的時(shí)間分辨熒光免疫檢測(cè)方法及試劑盒的制作方法
- 一種同時(shí)測(cè)定血漿中脂蛋白磷脂酶a2與c反應(yīng)蛋白含量的檢測(cè)方法及試劑盒的制作方法
- 血漿脂蛋白磷脂酶a2活性檢測(cè)試劑盒及其檢測(cè)方法